

PYK2 senses calcium through a disordered dimerization and calmodulin-binding element
- PMID: 35945264
- PMCID: PMC9363500
- DOI: 10.1038/s42003-022-03760-8
PYK2 senses calcium through a disordered dimerization and calmodulin-binding element
Abstract
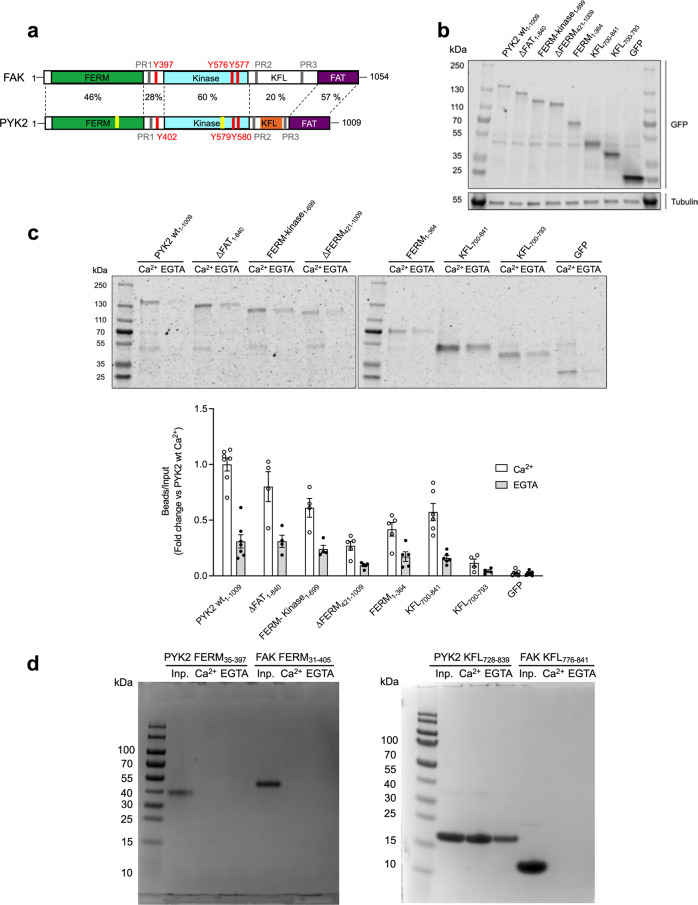
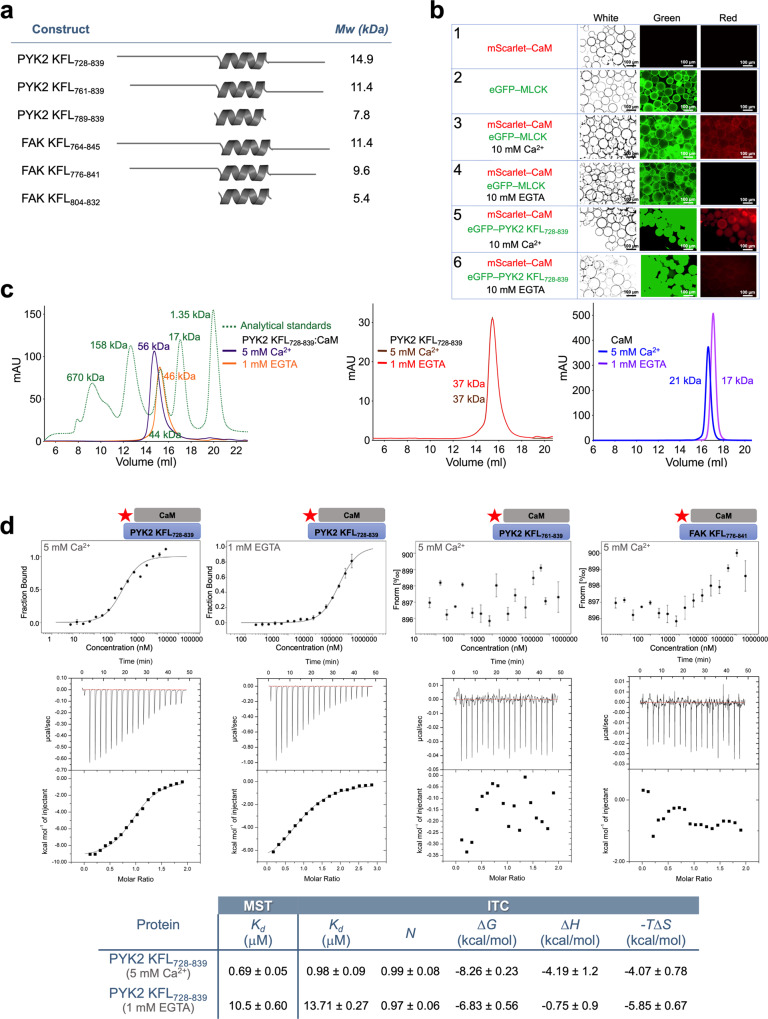
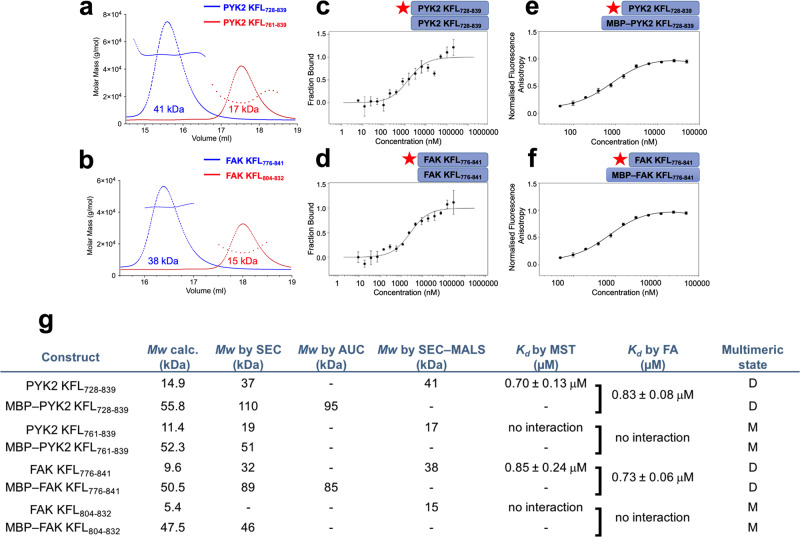
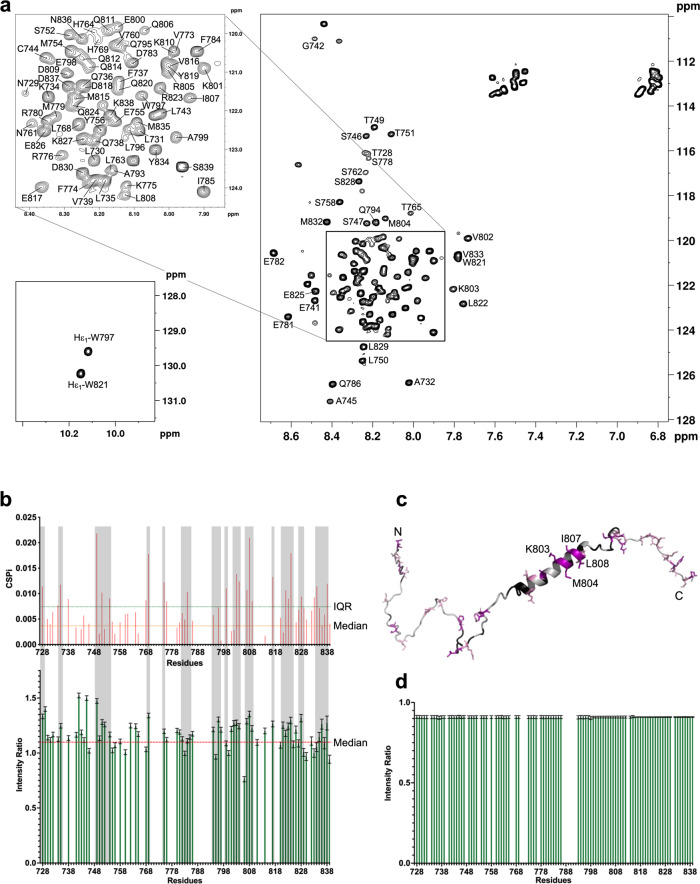
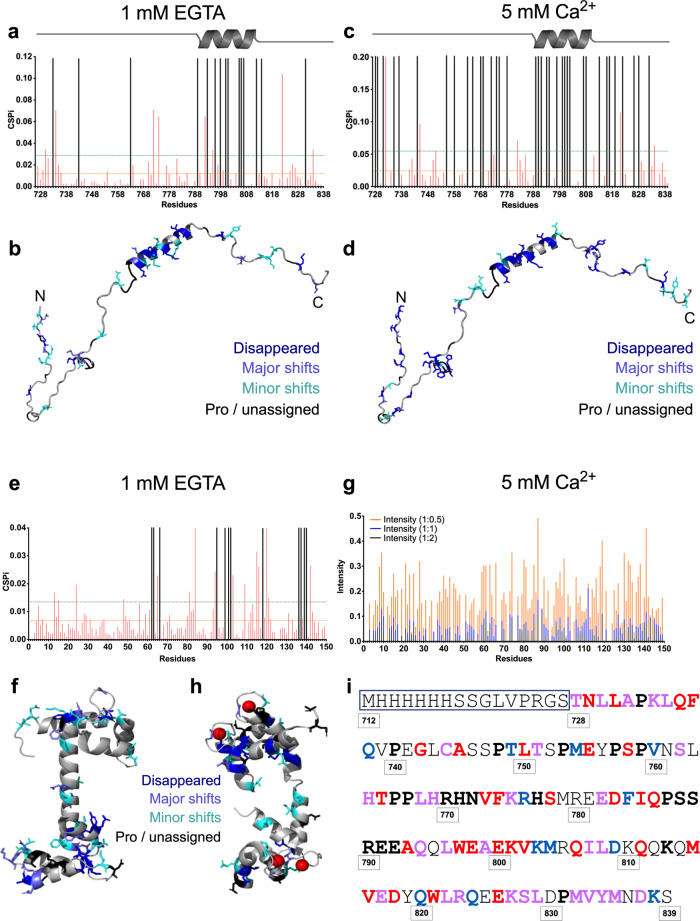
Multidomain kinases use many ways to integrate and process diverse stimuli. Here, we investigated the mechanism by which the protein tyrosine kinase 2-beta (PYK2) functions as a sensor and effector of cellular calcium influx. We show that the linker between the PYK2 kinase and FAT domains (KFL) encompasses an unusual calmodulin (CaM) binding element. PYK2 KFL is disordered and engages CaM through an ensemble of transient binding events. Calcium increases the association by promoting structural changes in CaM that expose auxiliary interaction opportunities. KFL also forms fuzzy dimers, and dimerization is enhanced by CaM binding. As a monomer, however, KFL associates with the PYK2 FERM-kinase fragment. Thus, we identify a mechanism whereby calcium influx can promote PYK2 self-association, and hence kinase-activating trans-autophosphorylation. Collectively, our findings describe a flexible protein module that expands the paradigms for CaM binding and self-association, and their use for controlling kinase activity.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

Similar articles
-
Protein-tyrosine kinase CAKbeta/PYK2 is activated by binding Ca2+/calmodulin to FERM F2 alpha2 helix and thus forming its dimer.Biochem J. 2008 Mar 15;410(3):513-23. doi: 10.1042/BJ20070665. Biochem J. 2008. PMID: 18031286
-
Analysis of the calcium-dependent regulation of proline-rich tyrosine kinase 2 by gonadotropin-releasing hormone.Mol Endocrinol. 2008 Oct;22(10):2322-35. doi: 10.1210/me.2008-0061. Epub 2008 Jul 17. Mol Endocrinol. 2008. PMID: 18635666 Free PMC article.
-
Calcium-dependent Pyk2 activation: a role for calmodulin?Biochem J. 2008 Mar 15;410(3):e3-4. doi: 10.1042/BJ20080133. Biochem J. 2008. PMID: 18290763
-
Conformation-Driven Phase Separation in the Linker Domain of Focal Adhesion Kinases.Biochemistry. 2025 Apr 15;64(8):1797-1806. doi: 10.1021/acs.biochem.4c00677. Epub 2025 Mar 26. Biochemistry. 2025. PMID: 40136251
-
The Pyk2 FERM regulates Pyk2 complex formation and phosphorylation.Cell Signal. 2011 Jan;23(1):288-96. doi: 10.1016/j.cellsig.2010.09.015. Epub 2010 Sep 16. Cell Signal. 2011. PMID: 20849950 Free PMC article.
Cited by
-
Molecularly imprinted nanoparticles reveal regulatory scaffolding features in Pyk2 tyrosine kinase.RSC Chem Biol. 2024 Mar 13;5(5):447-453. doi: 10.1039/d3cb00228d. eCollection 2024 May 8. RSC Chem Biol. 2024. PMID: 38725907 Free PMC article.
-
Exploring the evolving function of soluble intercellular adhesion molecule-1 in junction dynamics during spermatogenesis.Front Endocrinol (Lausanne). 2024 Jan 8;14:1281812. doi: 10.3389/fendo.2023.1281812. eCollection 2023. Front Endocrinol (Lausanne). 2024. PMID: 38260159 Free PMC article. Review.
-
Activated PyK2 and Its Associated Molecules Transduce Cellular Signaling from the Cancerous Milieu for Cancer Metastasis.Int J Mol Sci. 2022 Dec 7;23(24):15475. doi: 10.3390/ijms232415475. Int J Mol Sci. 2022. PMID: 36555115 Free PMC article. Review.
-
The Role of Pyk2 Kinase in Glioblastoma Progression and Therapeutic Targeting.Cancers (Basel). 2025 Aug 9;17(16):2611. doi: 10.3390/cancers17162611. Cancers (Basel). 2025. PMID: 40867240 Free PMC article. Review.
-
Pyk2 regulates sepsis-induced lung injury via ferroptosis.Iran J Basic Med Sci. 2023;26(11):1283-1290. doi: 10.22038/IJBMS.2023.69578.15153. Iran J Basic Med Sci. 2023. PMID: 37886006 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
