

mGluR5 PAMs rescue cortical and behavioural defects in a mouse model of CDKL5 deficiency disorder
- PMID: 35945276
- PMCID: PMC10156697
- DOI: 10.1038/s41386-022-01412-3
mGluR5 PAMs rescue cortical and behavioural defects in a mouse model of CDKL5 deficiency disorder
Abstract
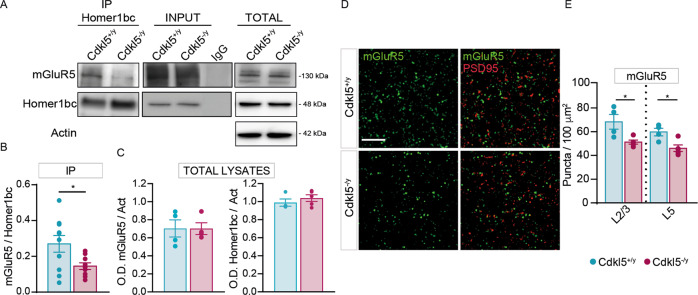
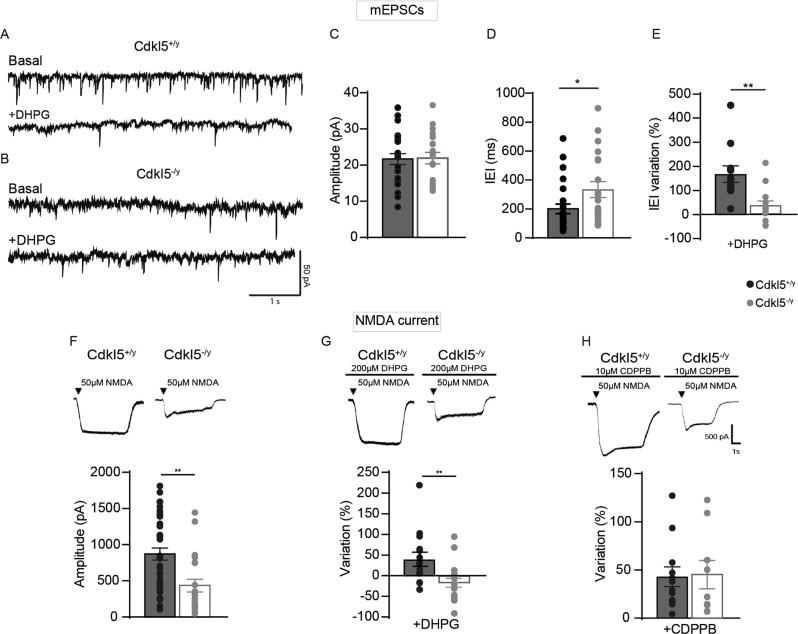
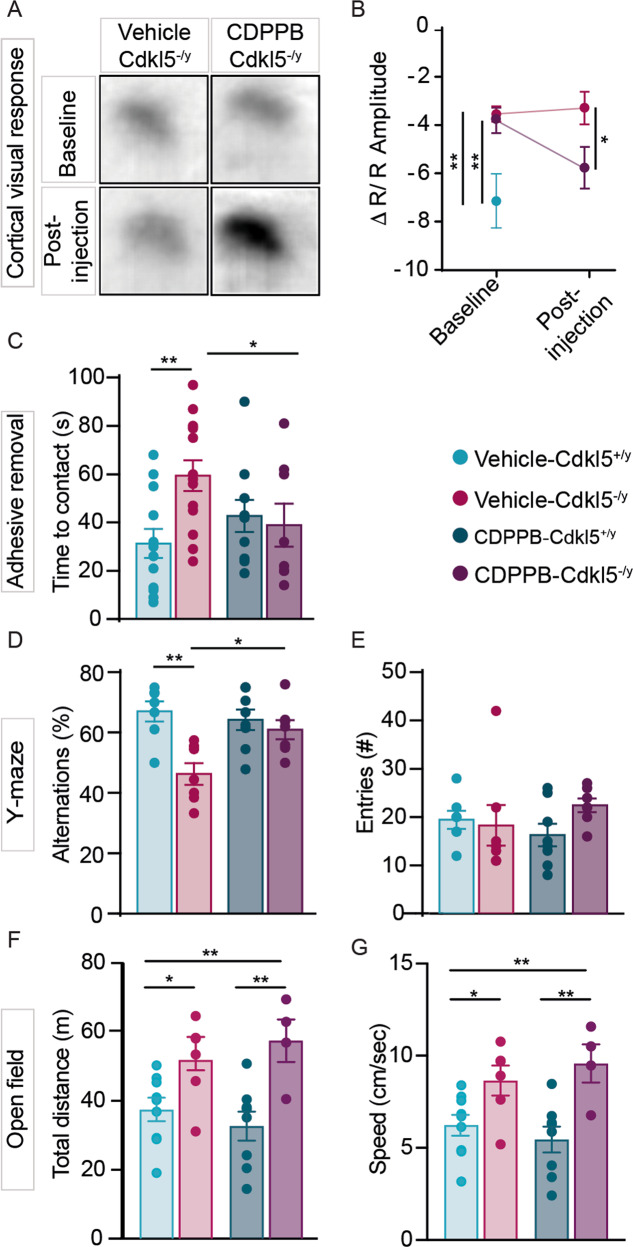
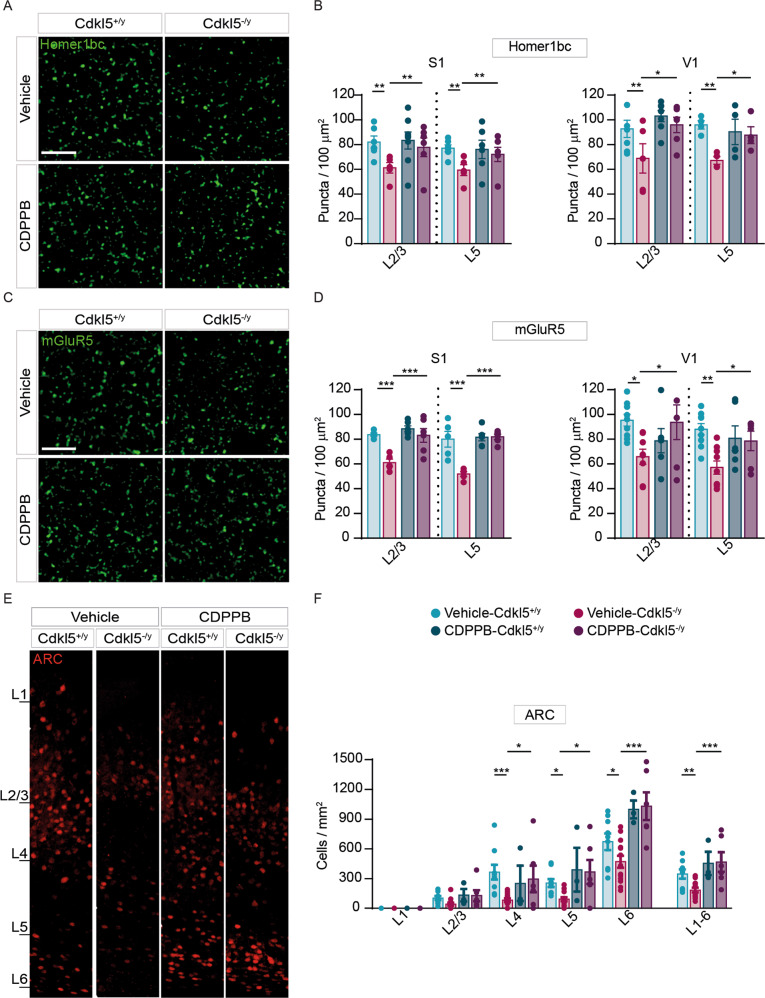
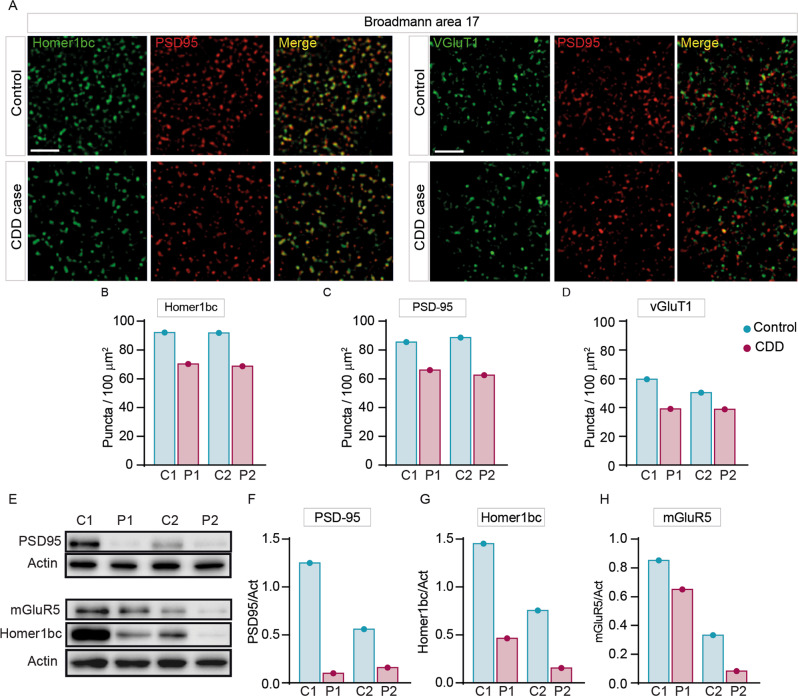
Cyclin-dependent kinase-like 5 (CDKL5) deficiency disorder (CDD) is a devastating rare neurodevelopmental disease without a cure, caused by mutations of the serine/threonine kinase CDKL5 highly expressed in the forebrain. CDD is characterized by early-onset seizures, severe intellectual disabilities, autistic-like traits, sensorimotor and cortical visual impairments (CVI). The lack of an effective therapeutic strategy for CDD urgently demands the identification of novel druggable targets potentially relevant for CDD pathophysiology. To this aim, we studied Class I metabotropic glutamate receptors 5 (mGluR5) because of their important role in the neuropathological signs produced by the lack of CDKL5 in-vivo, such as defective synaptogenesis, dendritic spines formation/maturation, synaptic transmission and plasticity. Importantly, mGluR5 function strictly depends on the correct expression of the postsynaptic protein Homer1bc that we previously found atypical in the cerebral cortex of Cdkl5-/y mice. In this study, we reveal that CDKL5 loss tampers with (i) the binding strength of Homer1bc-mGluR5 complexes, (ii) the synaptic localization of mGluR5 and (iii) the mGluR5-mediated enhancement of NMDA-induced neuronal responses. Importantly, we showed that the stimulation of mGluR5 activity by administering in mice specific positive-allosteric-modulators (PAMs), i.e., 3-Cyano-N-(1,3-diphenyl-1H-pyrazol-5-yl)benzamide (CDPPB) or RO6807794, corrected the synaptic, functional and behavioral defects shown by Cdkl5-/y mice. Notably, in the visual cortex of 2 CDD patients we found changes in synaptic organization that recapitulate those of mutant CDKL5 mice, including the reduced expression of mGluR5, suggesting that these receptors represent a promising therapeutic target for CDD.
© 2022. The Author(s), under exclusive licence to American College of Neuropsychopharmacology.
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Rusconi L, Salvatoni L, Giudici L, Bertani I, Kilstrup-Nielsen C, Broccoli V, et al. CDKL5 expression is modulated during neuronal development and its subcellular distribution is tightly regulated by the C-terminal tail. J Biol Chem. 2008;283:30101–11. doi: 10.1074/jbc.M804613200. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
