

Host translesion polymerases are required for viral genome integrity
- PMID: 35947614
- PMCID: PMC9388158
- DOI: 10.1073/pnas.2203203119
Host translesion polymerases are required for viral genome integrity
Abstract
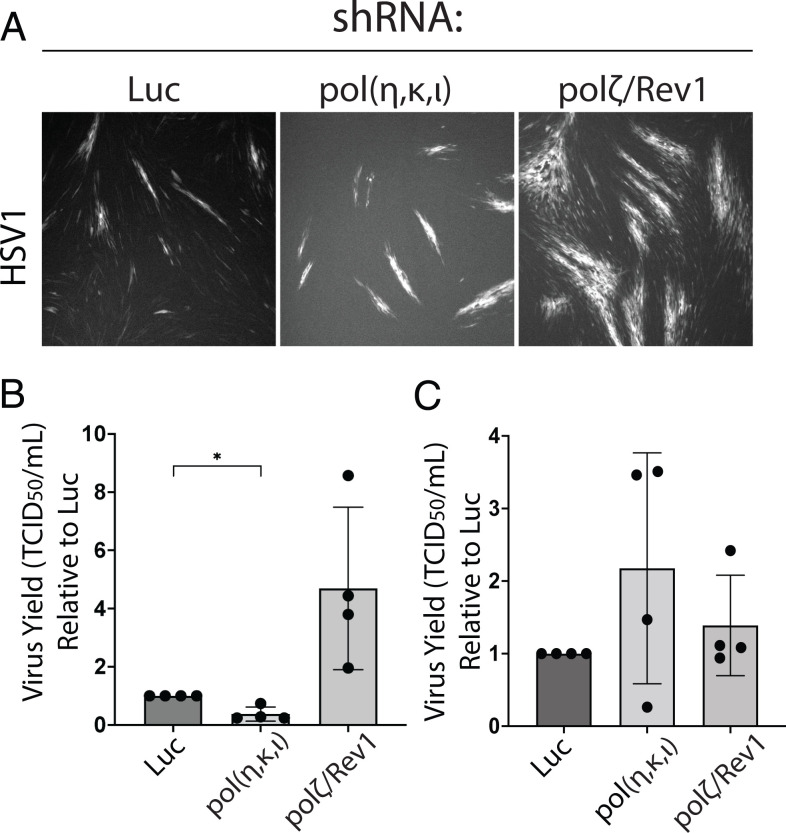
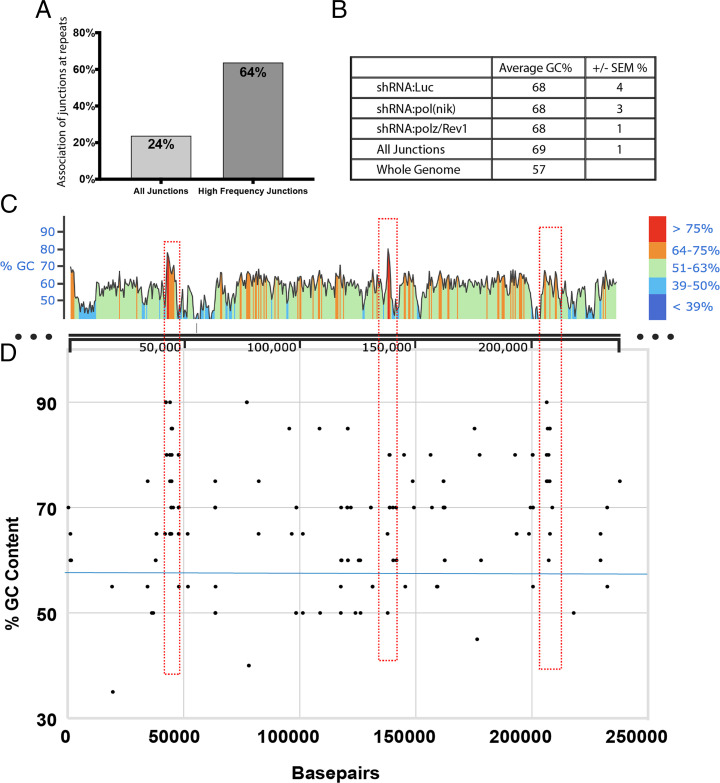
Human cells encode up to 15 DNA polymerases with specialized functions in chromosomal DNA synthesis and damage repair. In contrast, complex DNA viruses, such as those of the herpesviridae family, encode a single B-family DNA polymerase. This disparity raises the possibility that DNA viruses may rely on host polymerases for synthesis through complex DNA geometries. We tested the importance of error-prone Y-family polymerases involved in translesion synthesis (TLS) to human cytomegalovirus (HCMV) infection. We find most Y-family polymerases involved in the nucleotide insertion and bypass of lesions restrict HCMV genome synthesis and replication. In contrast, other TLS polymerases, such as the polymerase ζ complex, which extends past lesions, was required for optimal genome synthesis and replication. Depletion of either the polζ complex or the suite of insertion polymerases demonstrate that TLS polymerases suppress the frequency of viral genome rearrangements, particularly at GC-rich sites and repeat sequences. Moreover, while distinct from HCMV, replication of the related herpes simplex virus type 1 is impacted by host TLS polymerases, suggesting a broader requirement for host polymerases for DNA virus replication. These findings reveal an unexpected role for host DNA polymerases in ensuring viral genome stability.
Keywords: DNA repair; TLS polymerase; cytomegalovirus; genome replication; herpesvirus.
Conflict of interest statement
The authors declare no competing interest.
Figures




Similar articles
-
Complex roles for proliferating cell nuclear antigen in restricting human cytomegalovirus replication.mBio. 2025 May 14;16(5):e0045025. doi: 10.1128/mbio.00450-25. Epub 2025 Mar 25. mBio. 2025. PMID: 40130902 Free PMC article.
-
Complex roles for proliferating cell nuclear antigen in restricting human cytomegalovirus replication.bioRxiv [Preprint]. 2025 Jan 6:2025.01.06.631530. doi: 10.1101/2025.01.06.631530. bioRxiv. 2025. Update in: mBio. 2025 May 14;16(5):e0045025. doi: 10.1128/mbio.00450-25. PMID: 39829879 Free PMC article. Updated. Preprint.
-
Filling gaps in translesion DNA synthesis in human cells.Mutat Res Genet Toxicol Environ Mutagen. 2018 Dec;836(Pt B):127-142. doi: 10.1016/j.mrgentox.2018.02.004. Epub 2018 Feb 23. Mutat Res Genet Toxicol Environ Mutagen. 2018. PMID: 30442338 Review.
-
The Rev1-Polζ translesion synthesis mutasome: Structure, interactions and inhibition.Enzymes. 2019;45:139-181. doi: 10.1016/bs.enz.2019.07.001. Epub 2019 Aug 9. Enzymes. 2019. PMID: 31627876 Free PMC article. Review.
-
The roles of DNA polymerase ζ and the Y family DNA polymerases in promoting or preventing genome instability.Mutat Res. 2013 Mar-Apr;743-744:97-110. doi: 10.1016/j.mrfmmm.2012.11.002. Epub 2012 Nov 26. Mutat Res. 2013. PMID: 23195997 Free PMC article. Review.
Cited by
-
Human cytomegalovirus UL138 interaction with USP1 activates STAT1 in infection.PLoS Pathog. 2023 Jun 8;19(6):e1011185. doi: 10.1371/journal.ppat.1011185. eCollection 2023 Jun. PLoS Pathog. 2023. PMID: 37289831 Free PMC article.
-
Human Cytomegalovirus UL138 Interaction with USP1 Activates STAT1 in infection.bioRxiv [Preprint]. 2023 Feb 7:2023.02.07.527452. doi: 10.1101/2023.02.07.527452. bioRxiv. 2023. Update in: PLoS Pathog. 2023 Jun 8;19(6):e1011185. doi: 10.1371/journal.ppat.1011185. PMID: 36798153 Free PMC article. Updated. Preprint.
-
Evolutionary Dynamics of Accelerated Antiviral Resistance Development in Hypermutator Herpesvirus.Mol Biol Evol. 2024 Jul 3;41(7):msae119. doi: 10.1093/molbev/msae119. Mol Biol Evol. 2024. PMID: 38879872 Free PMC article.
-
Uracil-DNA Glycosylase of Murine Gammaherpesvirus 68 Binds Cognate Viral Replication Factors Independently of its Catalytic Residues.bioRxiv [Preprint]. 2023 May 19:2023.05.19.541466. doi: 10.1101/2023.05.19.541466. bioRxiv. 2023. Update in: mSphere. 2023 Oct 24;8(5):e0027823. doi: 10.1128/msphere.00278-23. PMID: 37398059 Free PMC article. Updated. Preprint.
-
Human TLS DNA polymerase: saviors or threats under replication stress?Mol Cell Biochem. 2025 May 23. doi: 10.1007/s11010-025-05291-2. Online ahead of print. Mol Cell Biochem. 2025. PMID: 40407973 Review.
References
-
- Ling H., Boudsocq F., Woodgate R., Yang W., Crystal structure of a Y-family DNA polymerase in action: A mechanism for error-prone and lesion-bypass replication. Cell 107, 91–102 (2001). - PubMed
-
- Sertic S., et al. , Coordinated activity of Y family TLS polymerases and EXO1 protects non-S phase cells from UV-induced cytotoxic lesions. Mol. Cell 70, 34–47.e4 (2018). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
