

Single base-pair resolution analysis of DNA binding motif with MoMotif reveals an oncogenic function of CTCF zinc-finger 1 mutation
- PMID: 35947648
- PMCID: PMC9410893
- DOI: 10.1093/nar/gkac658
Single base-pair resolution analysis of DNA binding motif with MoMotif reveals an oncogenic function of CTCF zinc-finger 1 mutation
Abstract
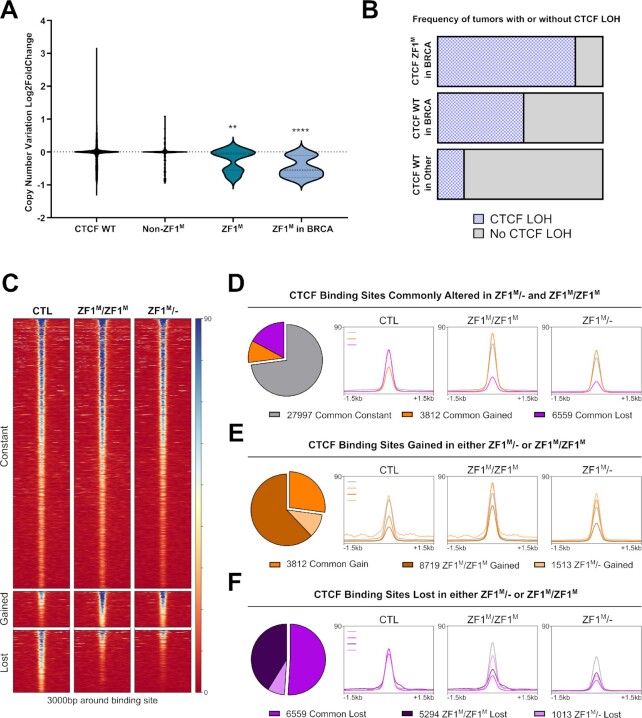
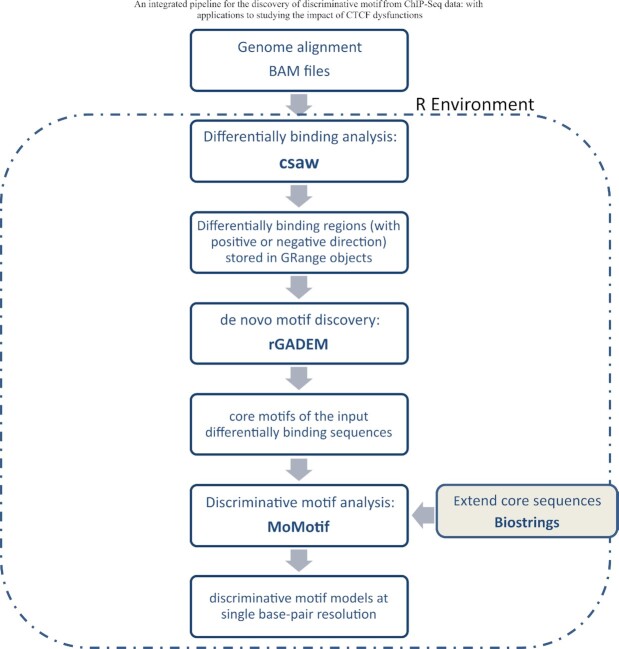
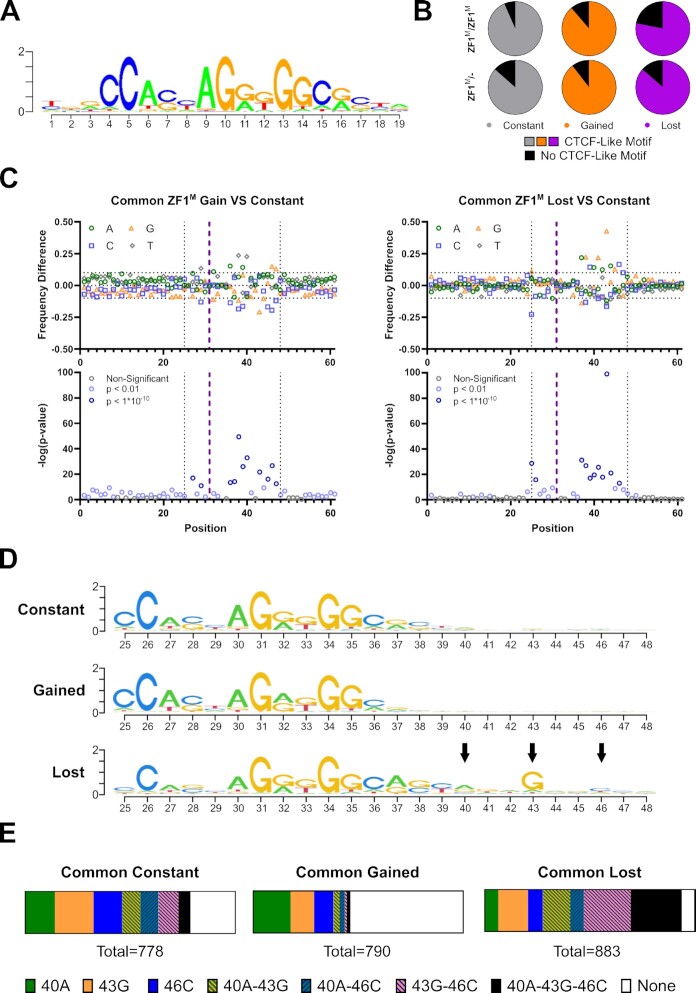
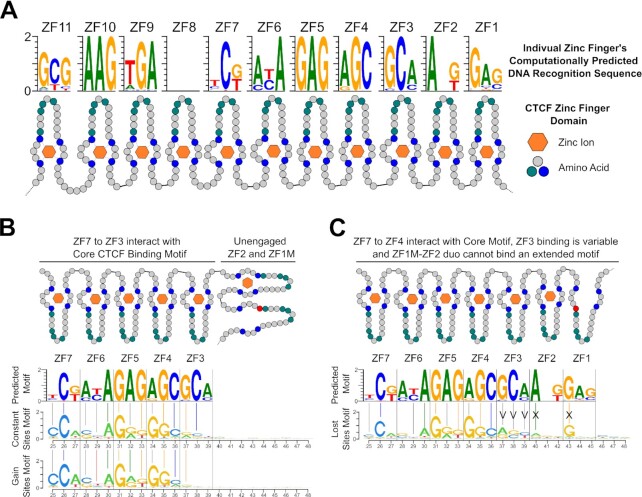
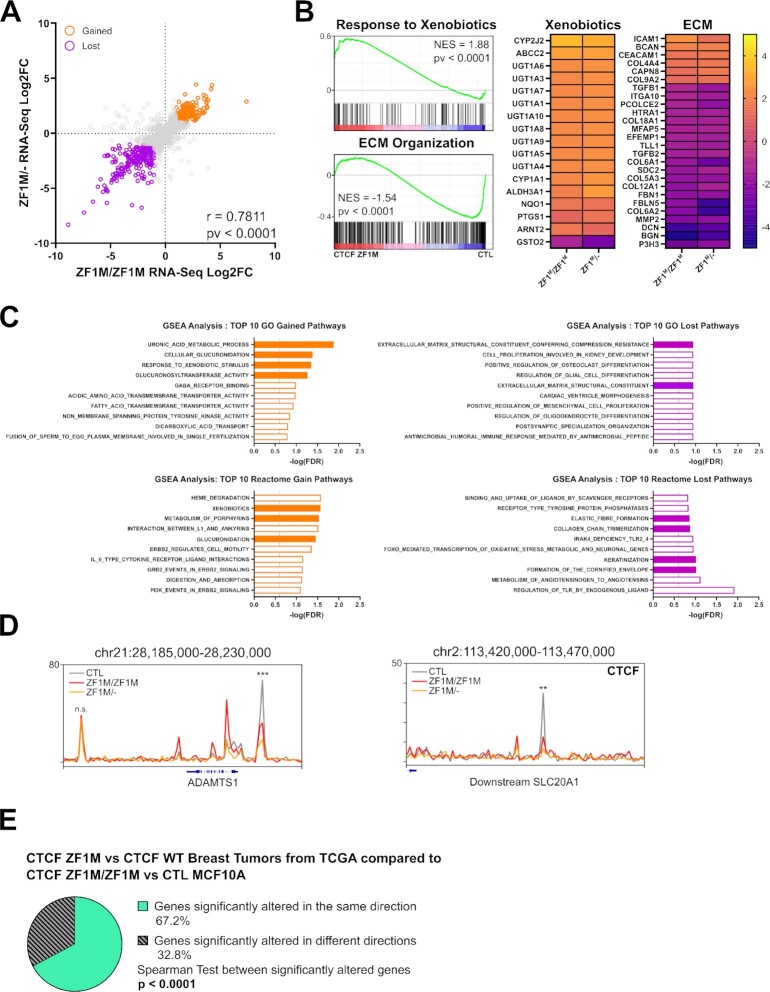
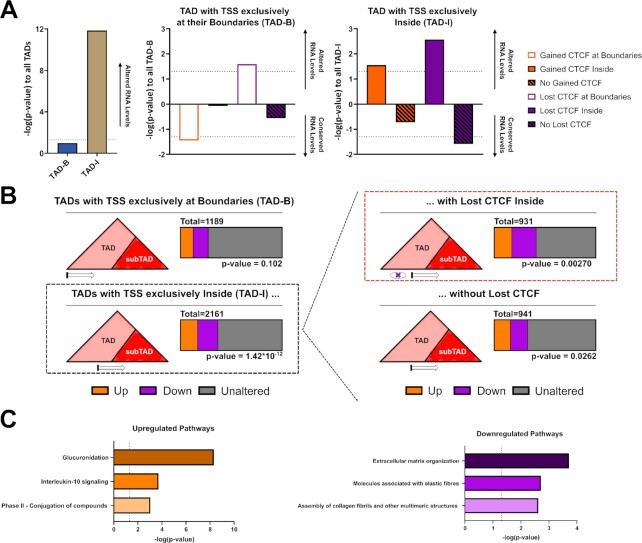
Defining the impact of missense mutations on the recognition of DNA motifs is highly dependent on bioinformatic tools that define DNA binding elements. However, classical motif analysis tools remain limited in their capacity to identify subtle changes in complex binding motifs between distinct conditions. To overcome this limitation, we developed a new tool, MoMotif, that facilitates a sensitive identification, at the single base-pair resolution, of complex, or subtle, alterations to core binding motifs, discerned from ChIP-seq data. We employed MoMotif to define the previously uncharacterized recognition motif of CTCF zinc-finger 1 (ZF1), and to further define the impact of CTCF ZF1 mutation on its association with chromatin. Mutations of CTCF ZF1 are exclusive to breast cancer and are associated with metastasis and therapeutic resistance, but the underlying mechanisms are unclear. Using MoMotif, we identified an extension of the CTCF core binding motif, necessitating a functional ZF1 to bind appropriately. Using a combination of ChIP-Seq and RNA-Seq, we discover that the inability to bind this extended motif drives an altered transcriptional program associated with the oncogenic phenotypes observed clinically. Our study demonstrates that MoMotif is a powerful new tool for comparative ChIP-seq analysis and characterising DNA-protein contacts.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
Similar articles
-
Neural network modeling of differential binding between wild-type and mutant CTCF reveals putative binding preferences for zinc fingers 1-2.BMC Genomics. 2022 Apr 12;23(1):295. doi: 10.1186/s12864-022-08486-9. BMC Genomics. 2022. PMID: 35410161 Free PMC article.
-
RNA Interactions Are Essential for CTCF-Mediated Genome Organization.Mol Cell. 2019 Nov 7;76(3):412-422.e5. doi: 10.1016/j.molcel.2019.08.015. Epub 2019 Sep 12. Mol Cell. 2019. PMID: 31522988 Free PMC article.
-
Structures of CTCF-DNA complexes including all 11 zinc fingers.Nucleic Acids Res. 2023 Sep 8;51(16):8447-8462. doi: 10.1093/nar/gkad594. Nucleic Acids Res. 2023. PMID: 37439339 Free PMC article.
-
CTCF: a Swiss-army knife for genome organization and transcription regulation.Essays Biochem. 2019 Apr 23;63(1):157-165. doi: 10.1042/EBC20180069. Print 2019 Apr 23. Essays Biochem. 2019. PMID: 30940740 Review.
-
Discovering a binary CTCF code with a little help from BORIS.Nucleus. 2018 Jan 1;9(1):33-41. doi: 10.1080/19491034.2017.1394536. Epub 2017 Dec 5. Nucleus. 2018. PMID: 29077515 Free PMC article. Review.
Cited by
-
A negatively charged region within carboxy-terminal domain maintains proper CTCF DNA binding.iScience. 2024 Nov 22;27(12):111452. doi: 10.1016/j.isci.2024.111452. eCollection 2024 Dec 20. iScience. 2024. PMID: 39720519 Free PMC article.
-
Repression of CADM1 transcription by HPV type 18 is mediated by three-dimensional rearrangement of promoter-enhancer interactions.PLoS Pathog. 2025 Jan 27;21(1):e1012506. doi: 10.1371/journal.ppat.1012506. eCollection 2025 Jan. PLoS Pathog. 2025. PMID: 39869645 Free PMC article.
-
Auxin-inducible degron 2 system deciphers functions of CTCF domains in transcriptional regulation.Genome Biol. 2023 Jan 26;24(1):14. doi: 10.1186/s13059-022-02843-3. Genome Biol. 2023. PMID: 36698211 Free PMC article.
-
Chromatin alternates between A and B compartments at kilobase scale for subgenic organization.Nat Commun. 2023 Jun 6;14(1):3303. doi: 10.1038/s41467-023-38429-1. Nat Commun. 2023. PMID: 37280210 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
