

Neuroblastoma suppressor of tumorigenicity 1 is a circulating protein associated with progression to end-stage kidney disease in diabetes
- PMID: 35947673
- PMCID: PMC9531292
- DOI: 10.1126/scitranslmed.abj2109
Neuroblastoma suppressor of tumorigenicity 1 is a circulating protein associated with progression to end-stage kidney disease in diabetes
Abstract
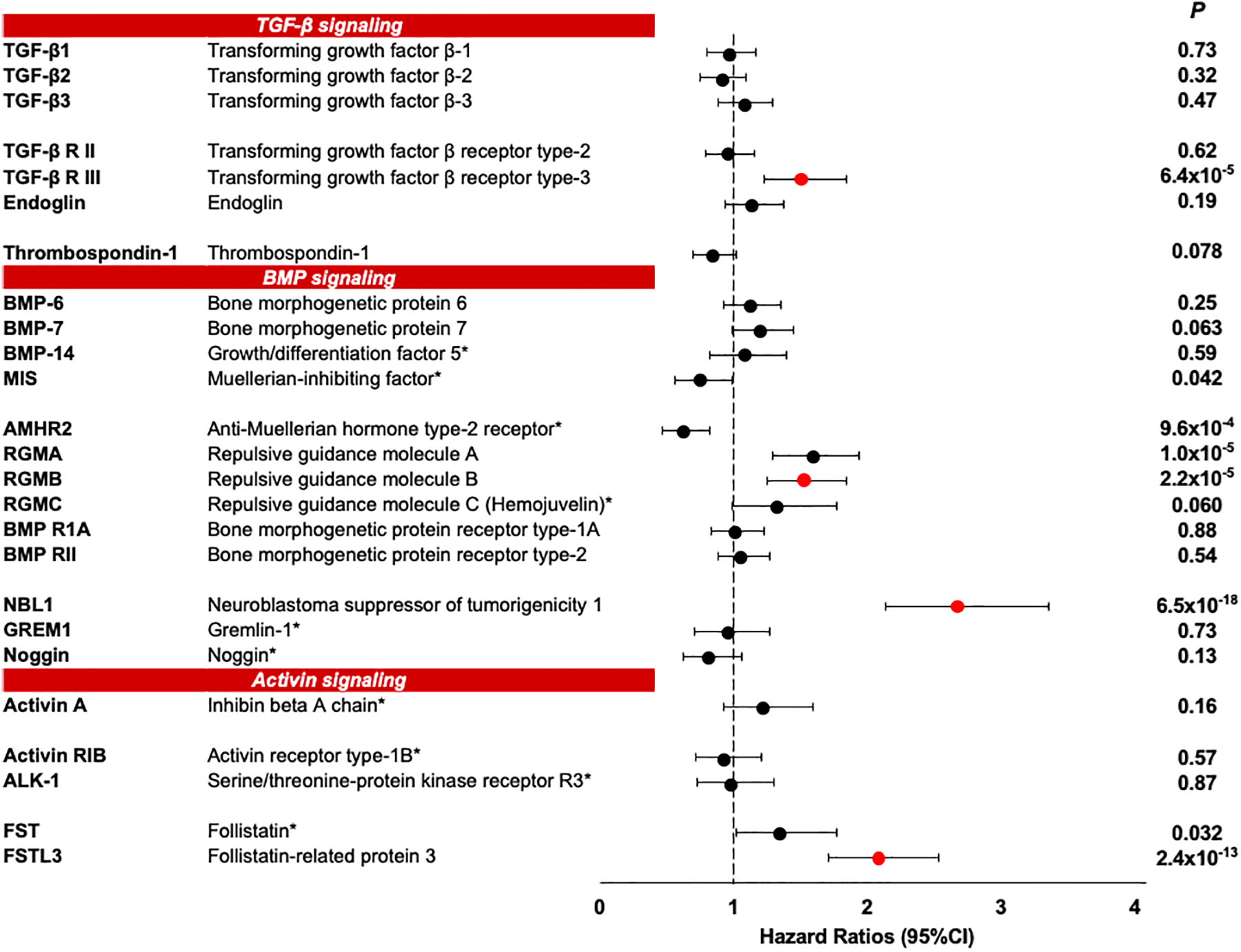
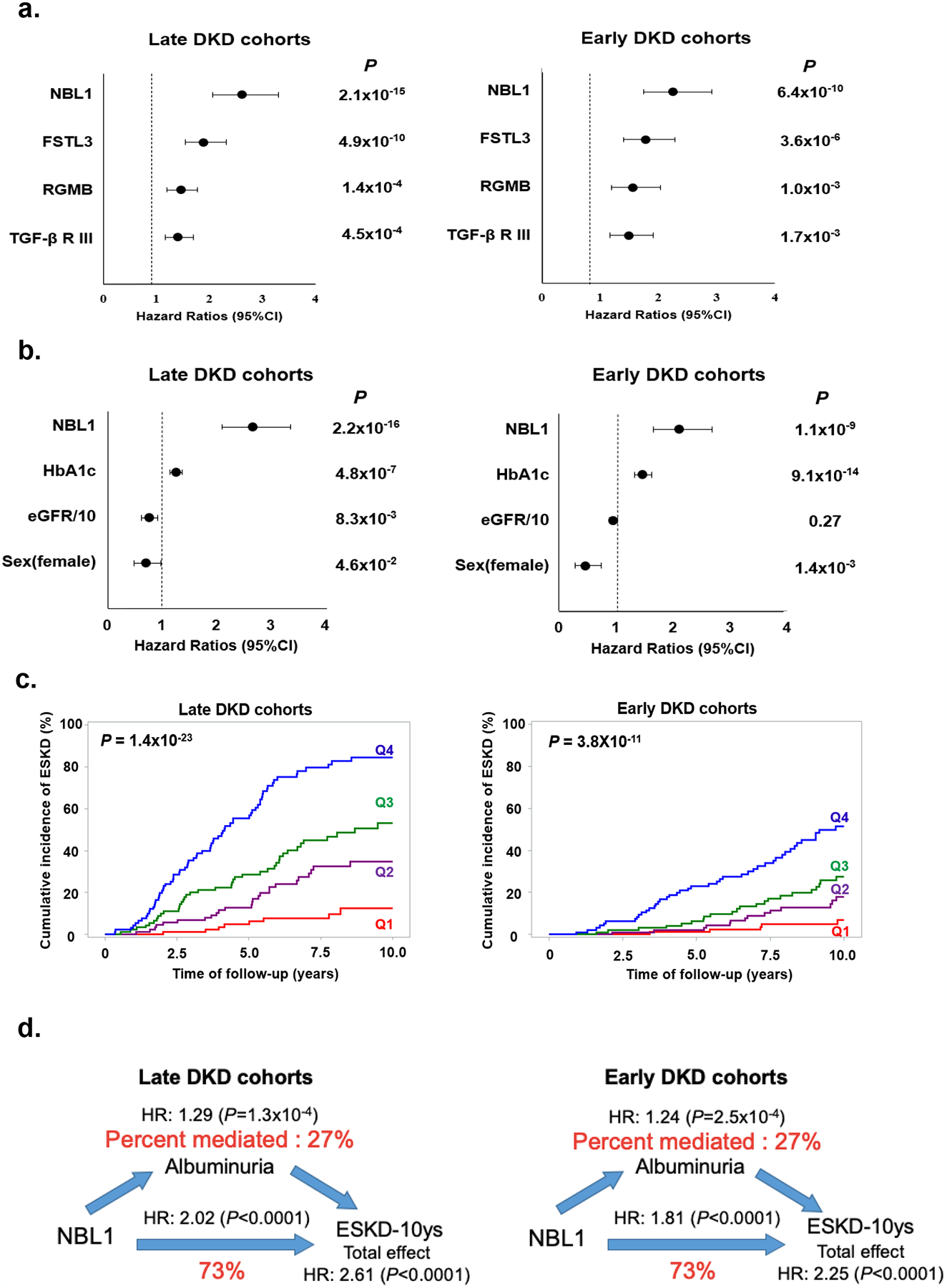
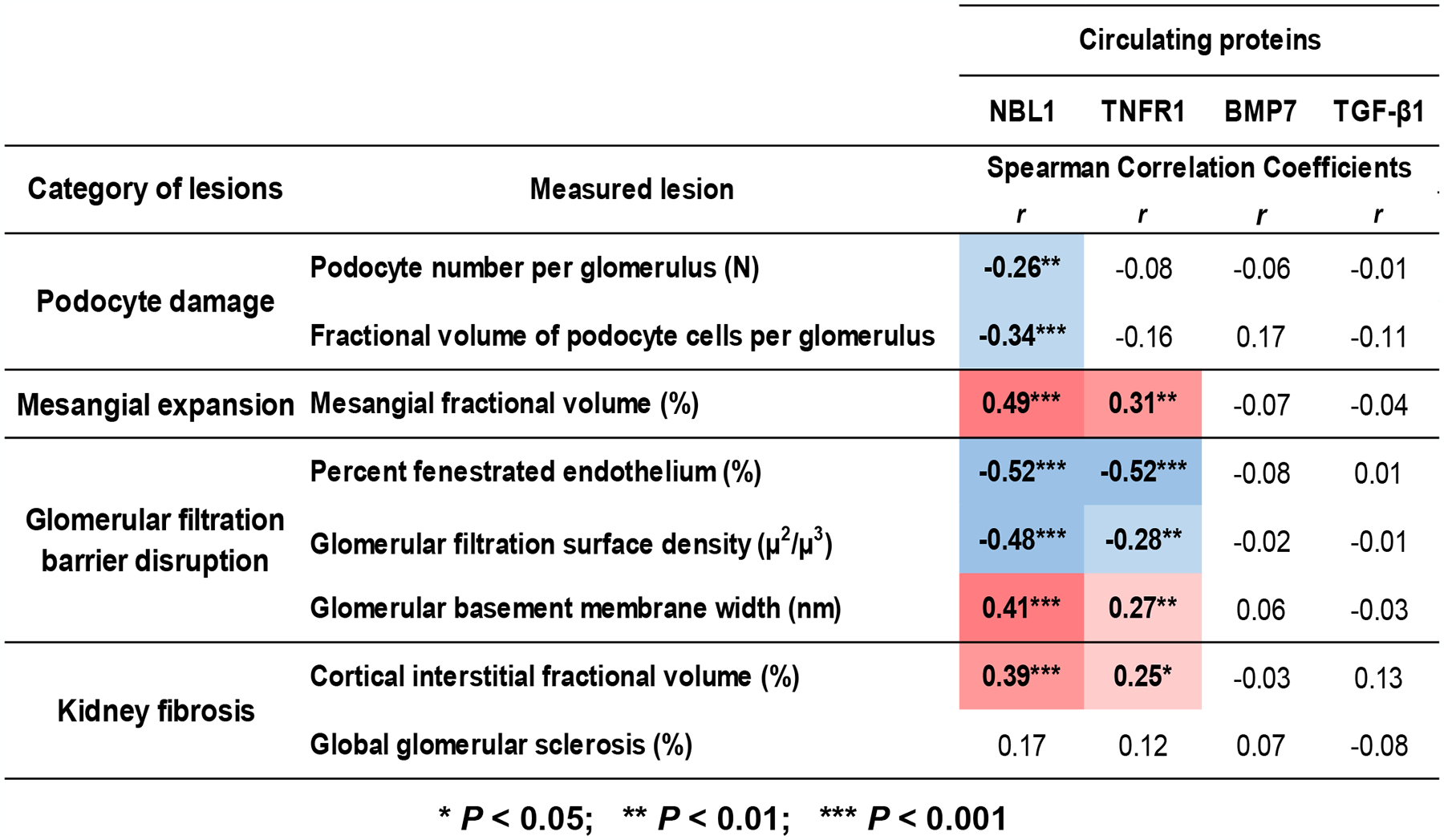
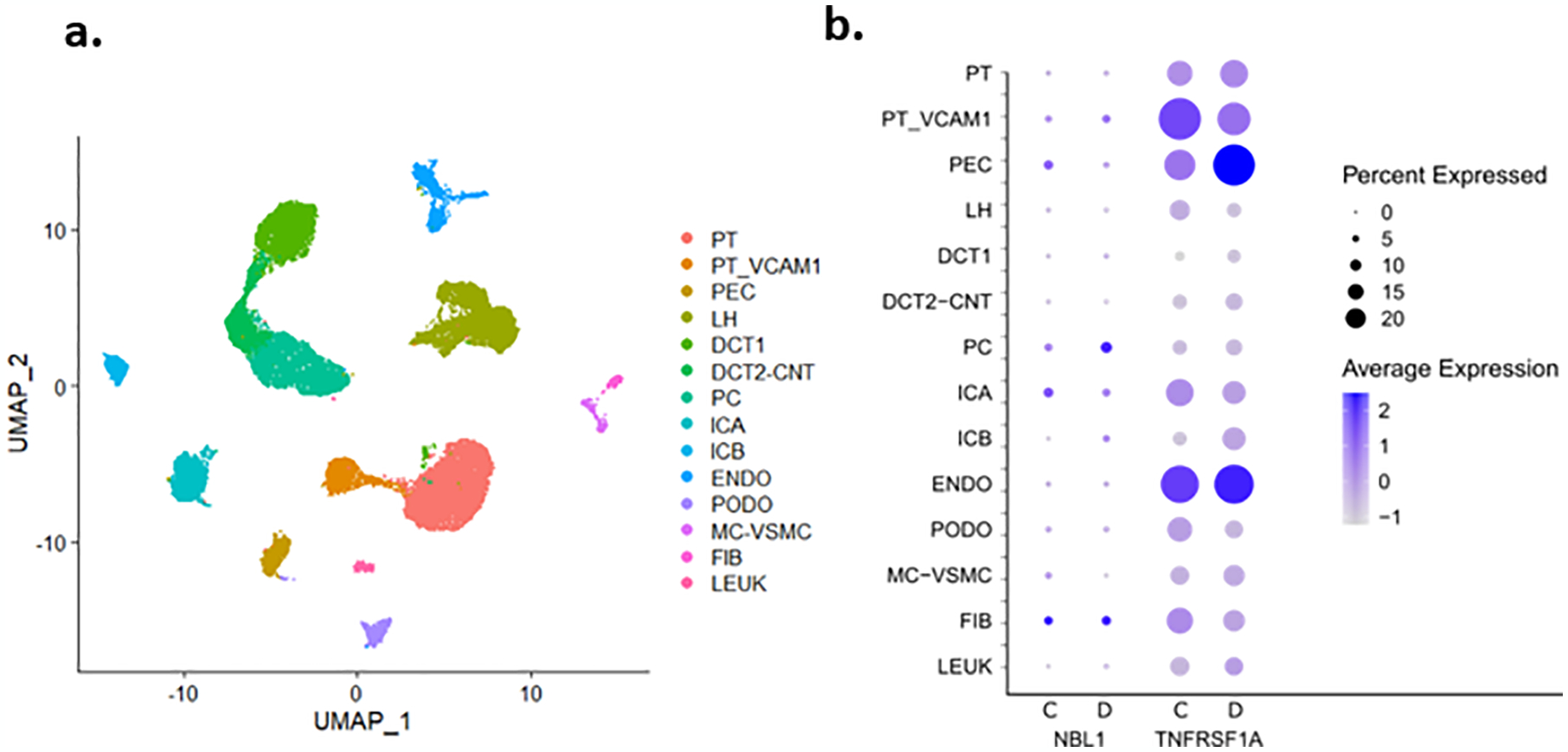
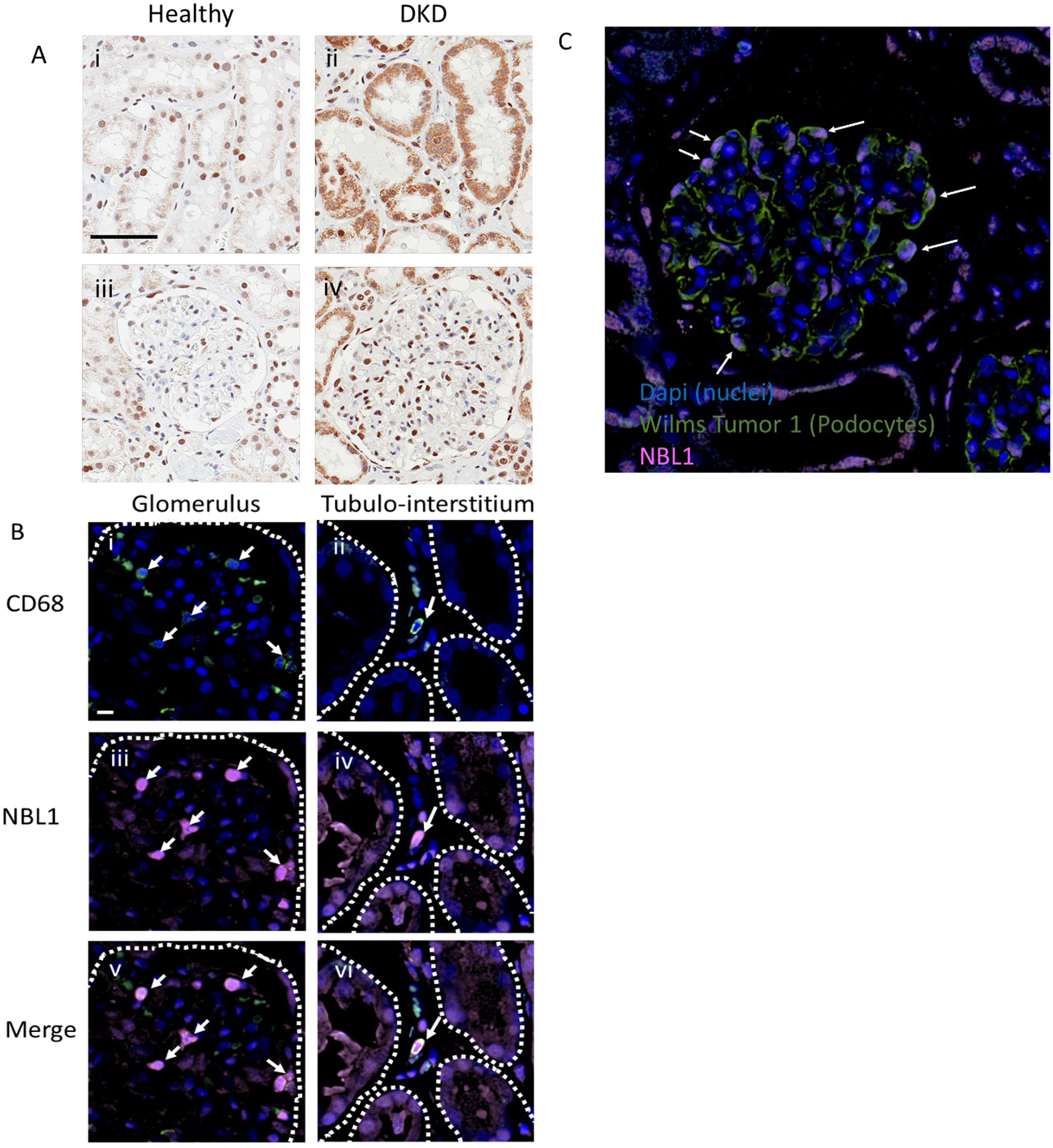
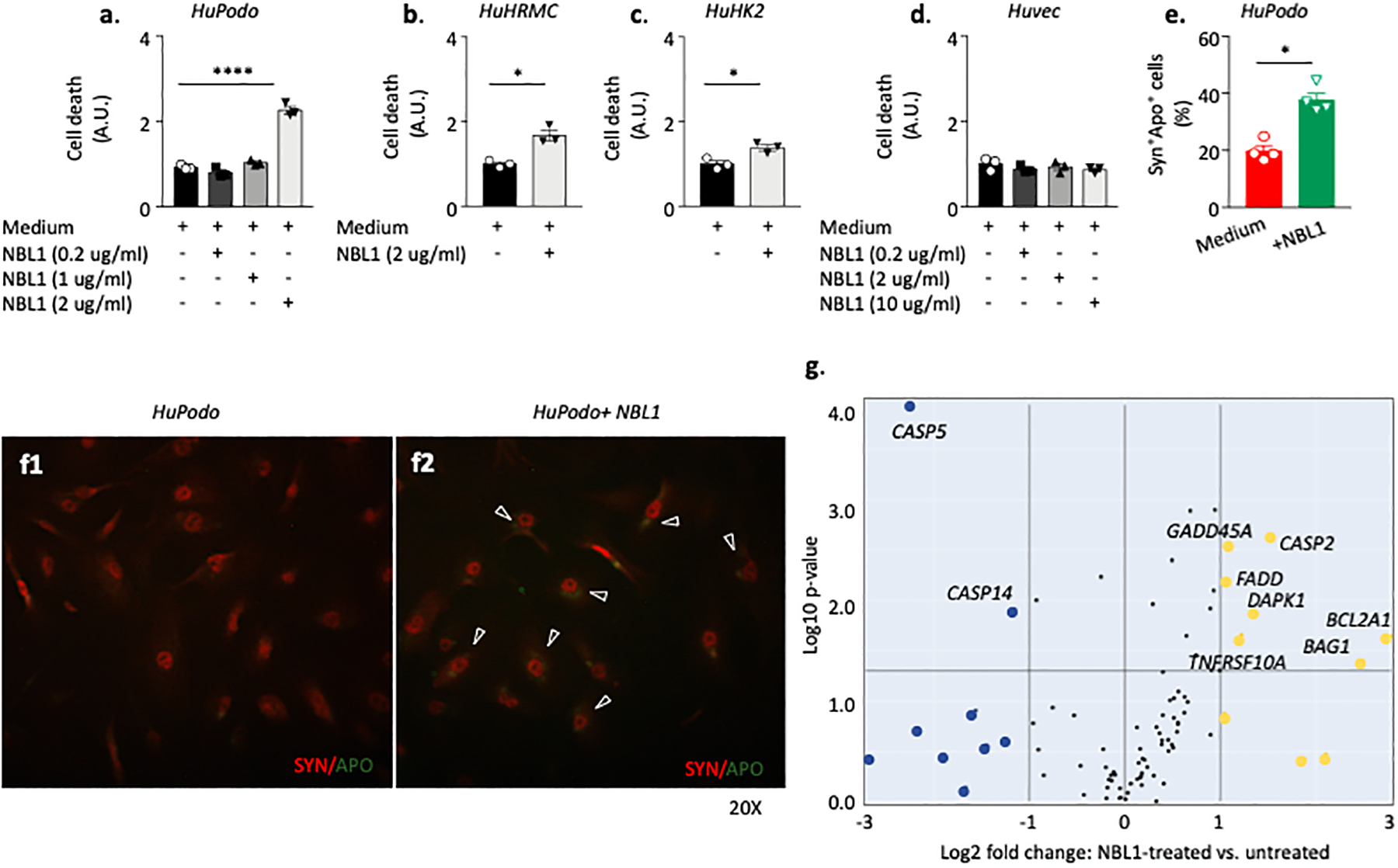
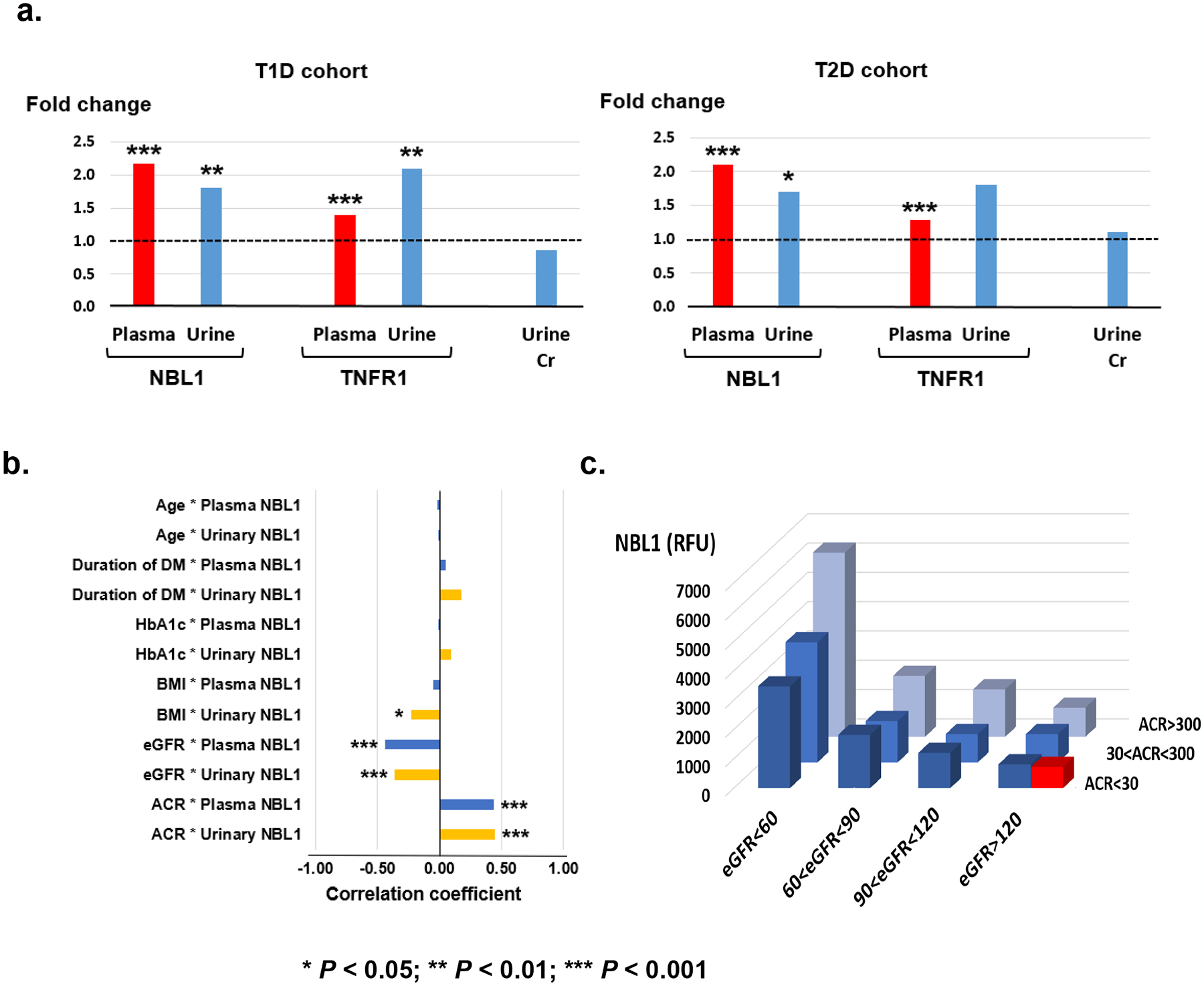
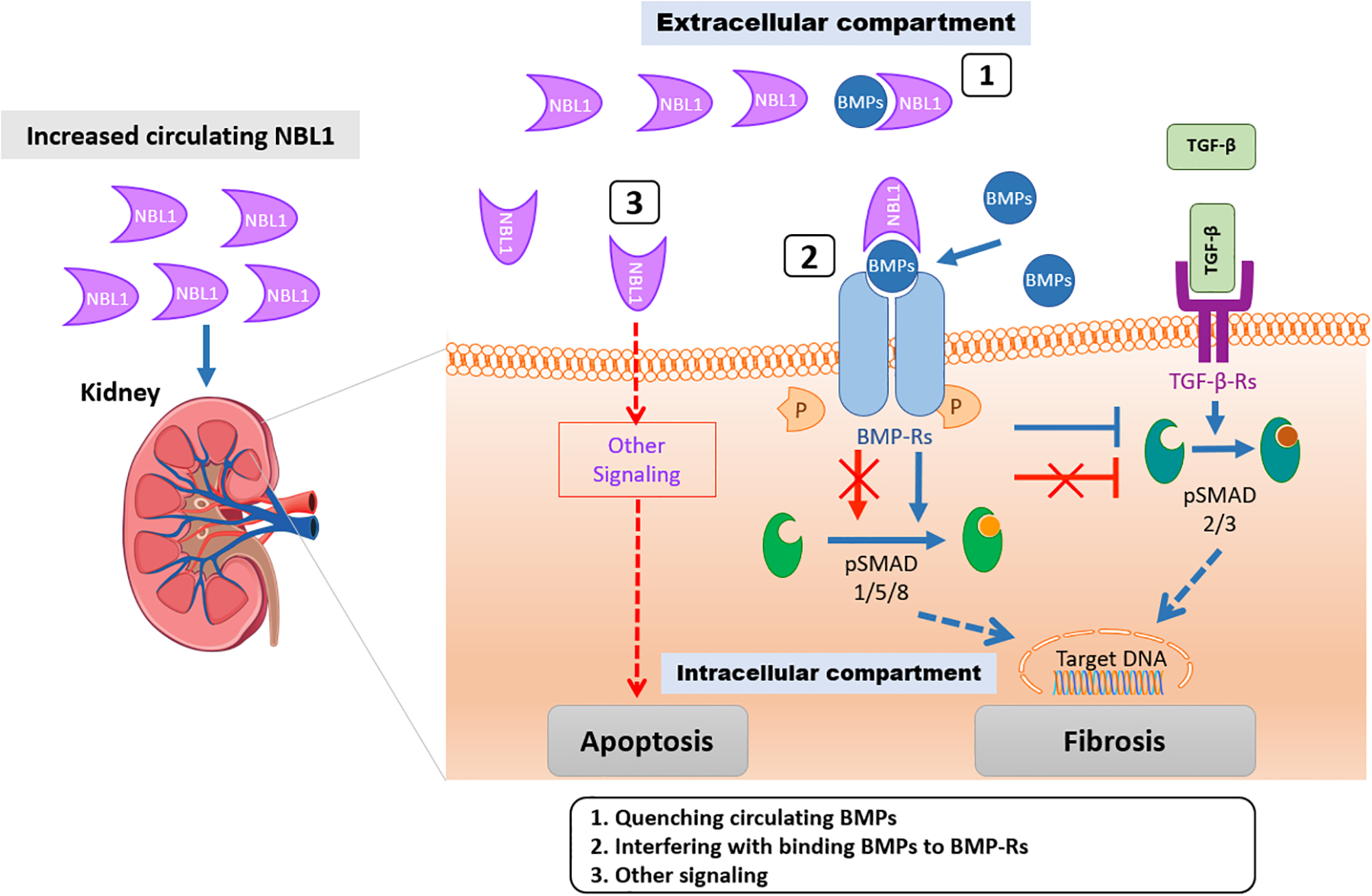
Circulating proteins associated with transforming growth factor-β (TGF-β) signaling are implicated in the development of diabetic kidney disease (DKD). It remains to be comprehensively examined which of these proteins are involved in the pathogenesis of DKD and its progression to end-stage kidney disease (ESKD) in humans. Using the SOMAscan proteomic platform, we measured concentrations of 25 TGF-β signaling family proteins in four different cohorts composed in total of 754 Caucasian or Pima Indian individuals with type 1 or type 2 diabetes. Of these 25 circulating proteins, we identified neuroblastoma suppressor of tumorigenicity 1 (NBL1, aliases DAN and DAND1), a small secreted protein known to inhibit members of the bone morphogenic protein family, to be most strongly and independently associated with progression to ESKD during 10-year follow-up in all cohorts. The extent of damage to podocytes and other glomerular structures measured morphometrically in 105 research kidney biopsies correlated strongly with circulating NBL1 concentrations. Also, in vitro exposure to NBL1 induced apoptosis in podocytes. In conclusion, circulating NBL1 may be involved in the disease process underlying progression to ESKD, and its concentration in circulation may identify subjects with diabetes at increased risk of progression to ESKD.
Figures
Comment in
-
Progress in the Early Detection of Risk of End-Stage Kidney Disease in Diabetes at Last?Clin Chem. 2023 Jul 5;69(7):673-675. doi: 10.1093/clinchem/hvac215. Clin Chem. 2023. PMID: 36762398 Free PMC article. No abstract available.
References
-
- Meng XM, Nikolic-Paterson DJ, Lan HY, TGF-β: the master regulator of fibrosis. Nat. Rev. Nephrol 12, 325–338 (2016). - PubMed
-
- Santibanez JF, Quintanilla M, Bernabeu C, TGF-beta/TGF-beta receptor system and its role in physiological and pathological conditions. Clin. Sci (Lond) 121, 233–251 (2011). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
