

Cancer Associated Fibroblast (CAF) Regulation of PDAC Parenchymal (CPC) and CSC Phenotypes Is Modulated by ECM Composition
- PMID: 35954400
- PMCID: PMC9367491
- DOI: 10.3390/cancers14153737
Cancer Associated Fibroblast (CAF) Regulation of PDAC Parenchymal (CPC) and CSC Phenotypes Is Modulated by ECM Composition
Abstract
Background: Pancreatic ductal adenocarcinoma (PDAC) is one of the deadliest of all cancers, having one of the lowest five-year survival rates. One of its hallmarks is a dense desmoplastic stroma consisting in the abnormal accumulation of extracellular matrix (ECM) components, especially Collagen I. This highly fibrotic stroma embeds the bulk cancer (parenchymal) cells (CPCs), cancer stem cells (CSCs) and the main producers of the stromal reaction, the Cancer Associated Fibroblasts (CAFs). Little is known about the role of the acellular ECM in the interplay of the CAFs with the different tumor cell types in determining their phenotypic plasticity and eventual cell fate.
Methods: Here, we analyzed the role of ECM collagen I in modulating the effect of CAF-derived signals by incubating PDAC CPCs and CSCs grown on ECM mimicking early (low collagen I levels) and late (high collagen I levels) stage PDAC stroma with conditioned medium from primary cultured CAFs derived from patients with PDAC in a previously described three-dimensional (3D) organotypic model of PDAC.
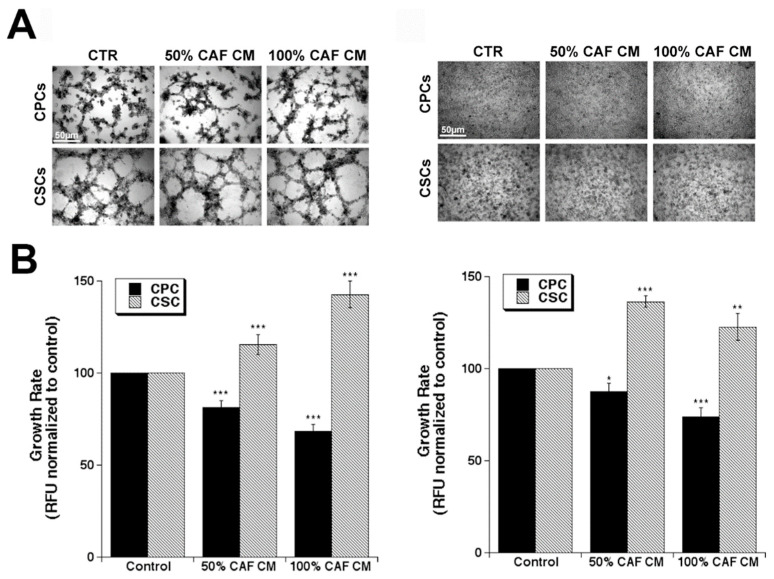
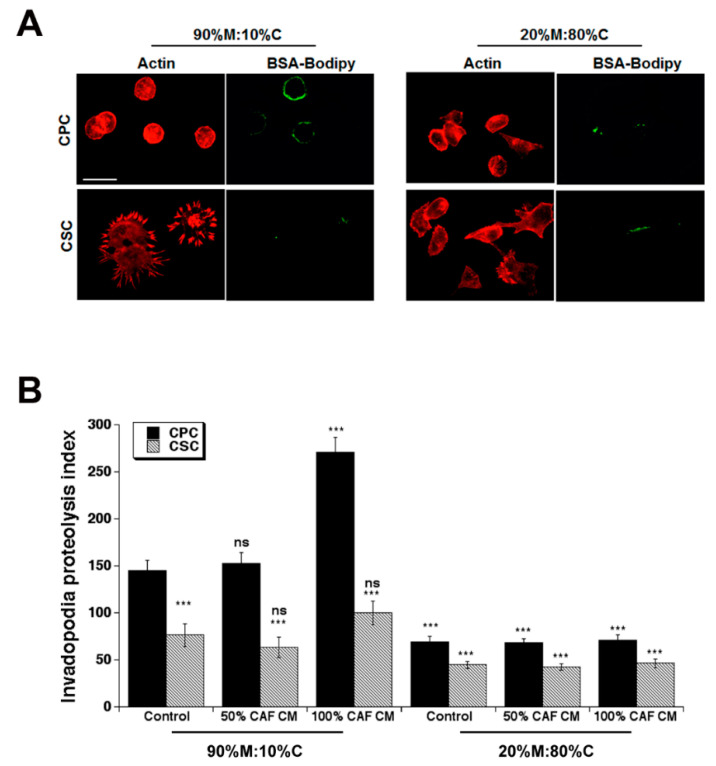
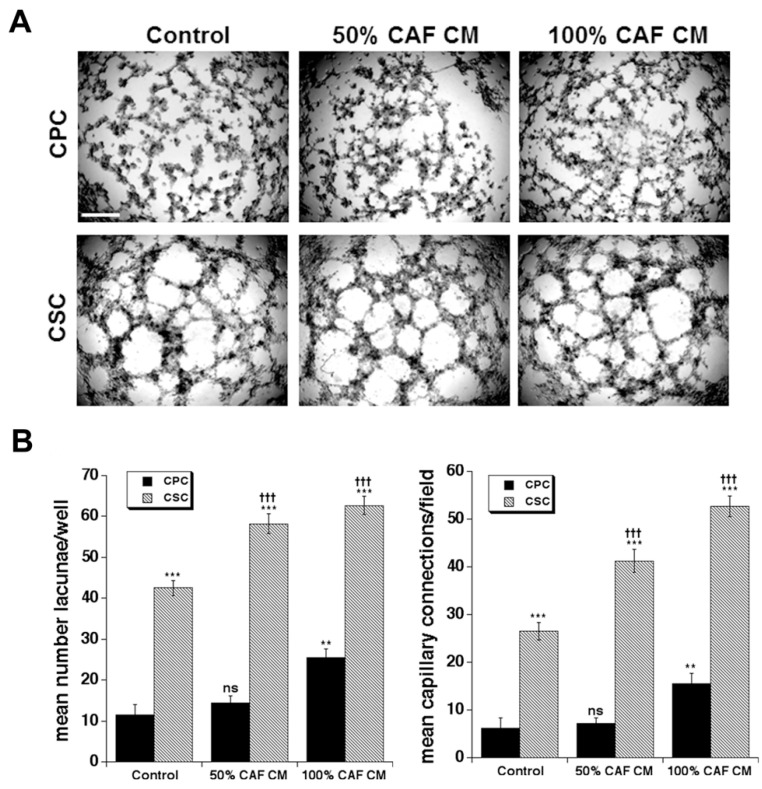
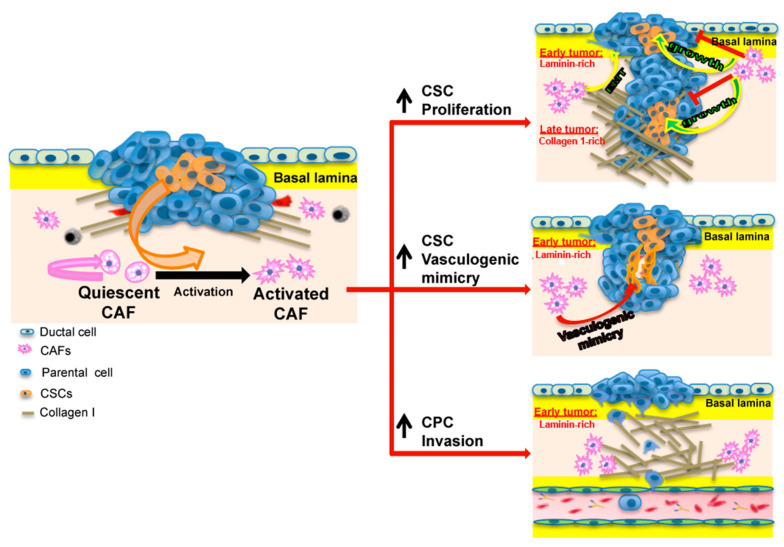
Results: We found that CAFs (1) reduced CPC growth while favoring CSC growth independently of the ECM; (2) increased the invasive capacity of only CPCs on the ECM mimicking the early tumor; and (3) favored vasculogenic mimicry (VM) especially of the CSCs on the ECM mimicking an early tumor.
Conclusions: We conclude that the CAFs and acellular stromal components interact to modulate the tumor behaviors of the PDAC CPC and CSC cell types and drive metastatic progression by stimulating the phenotypic characteristics of each tumor cell type that contribute to metastasis.
Keywords: 3D organotypic cultures; desmoplastic reaction; invadopodia; pancreatic ductal adenocarcinoma; vasculogenic mimicry.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript; or in the decision to publish the results.
Figures
References
-
- Biondani G., Zeeberg K., Greco M.R., Cannone S., Dando I., Pozza E.D., Mastrodonato M., Forciniti S., Casavola V., Palmieri M., et al. Extracellular Matrix composition modulates PDAC parenchymal and stem cell plasticity and behavior through the secretome. FEBS J. 2018;285:2104–2124. doi: 10.1111/febs.14471. - DOI - PubMed
Grants and funding
- 2013-00038330; 2015-00059283/Fondation de France
- PJA 20161204740/Fondation ARC pour la Recherche sur le Cancer
- GB/MA/IQ-10607/Ligue contre le Cancer
- No. 813834-pHioniC-H2020-MSCA-ITN-2018/Marie Skłodowska-Curie
- FP7-PEOPLE-2011-ITN Grant Agreement No. 289648/Marie Curie Initial Training Network IonTraC
LinkOut - more resources
Full Text Sources
