

Reg-1α, a New Substrate of Calpain-2 Depending on Its Glycosylation Status
- PMID: 35955718
- PMCID: PMC9369050
- DOI: 10.3390/ijms23158591
Reg-1α, a New Substrate of Calpain-2 Depending on Its Glycosylation Status
Abstract
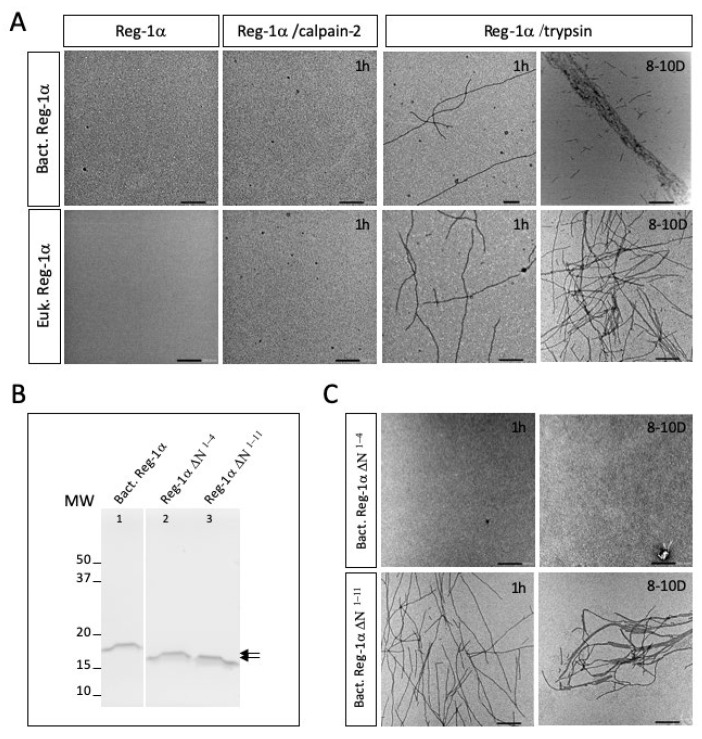
Reg-1α/lithostathine, a protein mainly associated with the digestive system, was previously shown to be overexpressed in the pre-clinical stages of Alzheimer's disease. In vitro, the glycosylated protein was reported to form fibrils at physiological pH following the proteolytic action of trypsin. However, the nature of the protease able to act in the central nervous system is unknown. In the present study, we showed that Reg-1α can be cleaved in vitro by calpain-2, the calcium activated neutral protease, overexpressed in neurodegenerative diseases. Using chemical crosslinking experiments, we found that the two proteins can interact with each other. Identification of the cleavage site using mass spectrometry, between Gln4 and Thr5, was found in agreement with the in silico prediction of the calpain cleavage site, in a position different from the one reported for trypsin, i.e., Arg11-Ile12 peptide bond. We showed that the cleavage was impeded by the presence of the neighboring glycosylation of Thr5. Moreover, in vitro studies using electron microscopy showed that calpain-cleaved protein does not form fibrils as observed after trypsin cleavage. Collectively, our results show that calpain-2 cleaves Reg-1α in vitro, and that this action is not associated with fibril formation.
Keywords: Reg-1α; calpain; cleavage; fibril; trypsin.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
Biophysical characterization of lithostathine. Evidences for a polymeric structure at physiological pH and a proteolysis mechanism leading to the formation of fibrils.J Biol Chem. 1999 Aug 6;274(32):22266-74. doi: 10.1074/jbc.274.32.22266. J Biol Chem. 1999. PMID: 10428794
-
Regenerating islet-derived 1α (Reg-1α) protein is new neuronal secreted factor that stimulates neurite outgrowth via exostosin Tumor-like 3 (EXTL3) receptor.J Biol Chem. 2012 Feb 10;287(7):4726-39. doi: 10.1074/jbc.M111.260349. Epub 2011 Dec 9. J Biol Chem. 2012. PMID: 22158612 Free PMC article.
-
Protein-X, Pancreatic Stone-, Pancreatic thread-, reg-protein, P19, lithostathine, and now what? Characterization, structural analysis and putative function(s) of the major non-enzymatic protein of pancreatic secretions.Curr Protein Pept Sci. 2001 Mar;2(1):19-42. doi: 10.2174/1389203013381233. Curr Protein Pept Sci. 2001. PMID: 12369899 Review.
-
Proteolytic processing of beta-amyloid precursor by calpain I.J Neurosci. 1990 Jul;10(7):2400-11. doi: 10.1523/JNEUROSCI.10-07-02400.1990. J Neurosci. 1990. PMID: 2115911 Free PMC article.
-
Digestive versus regulatory proteases: on calpain action in vivo.Biol Chem. 2005 Jul;386(7):609-12. doi: 10.1515/BC.2005.071. Biol Chem. 2005. PMID: 16207081 Review.
Cited by
-
Controversial Roles of Regenerating Family Proteins in Tissue Repair and Tumor Development.Biomedicines. 2024 Dec 26;13(1):24. doi: 10.3390/biomedicines13010024. Biomedicines. 2024. PMID: 39857608 Free PMC article. Review.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
