

Plant_SNP_TATA_Z-Tester: A Web Service That Unequivocally Estimates the Impact of Proximal Promoter Mutations on Plant Gene Expression
- PMID: 35955817
- PMCID: PMC9369029
- DOI: 10.3390/ijms23158684
Plant_SNP_TATA_Z-Tester: A Web Service That Unequivocally Estimates the Impact of Proximal Promoter Mutations on Plant Gene Expression
Abstract
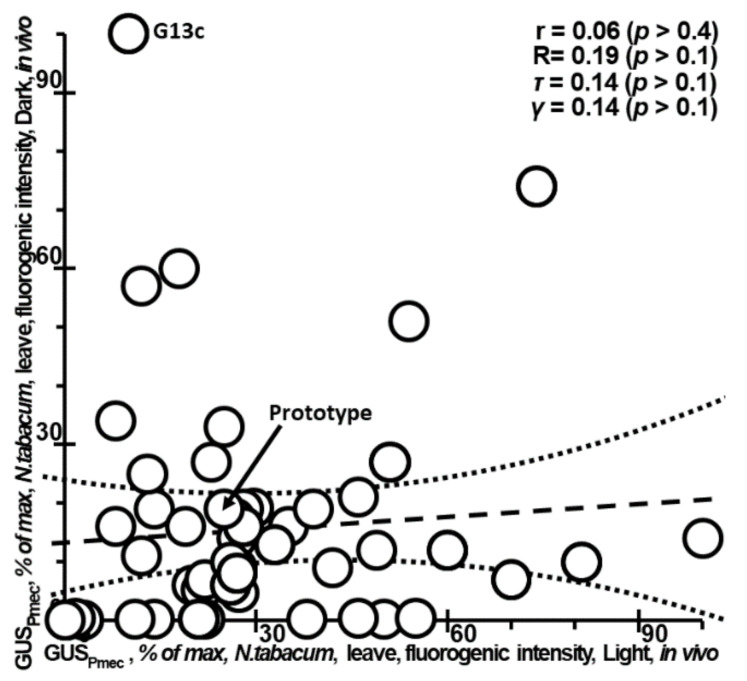
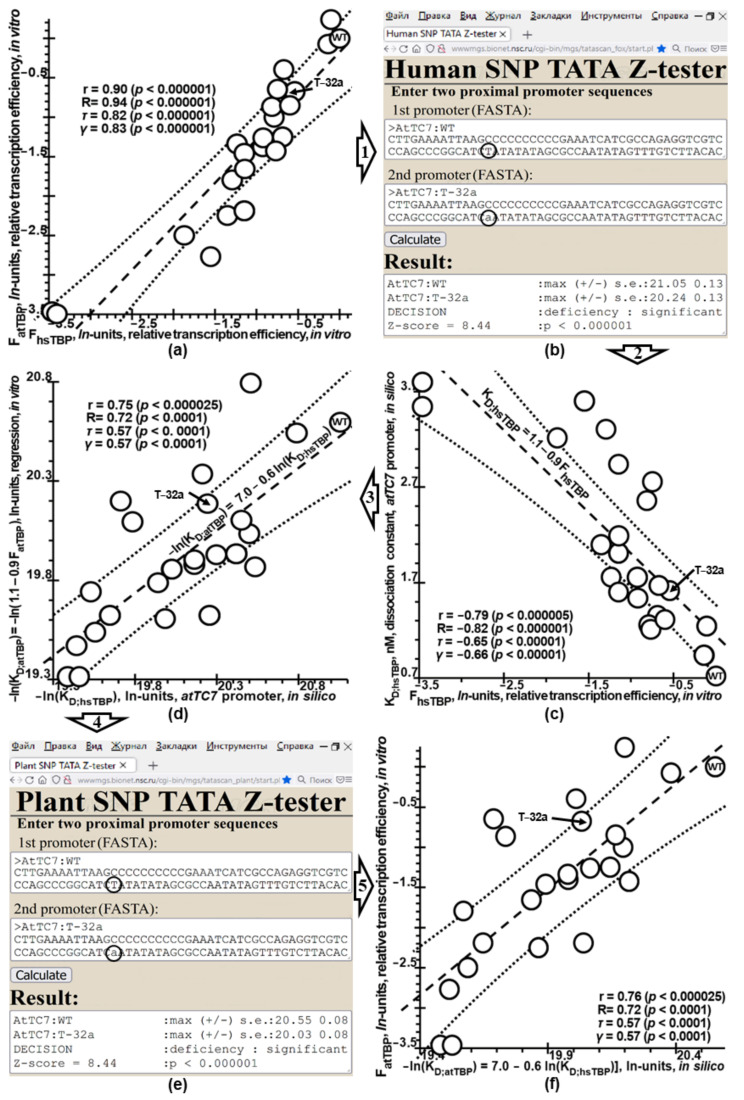
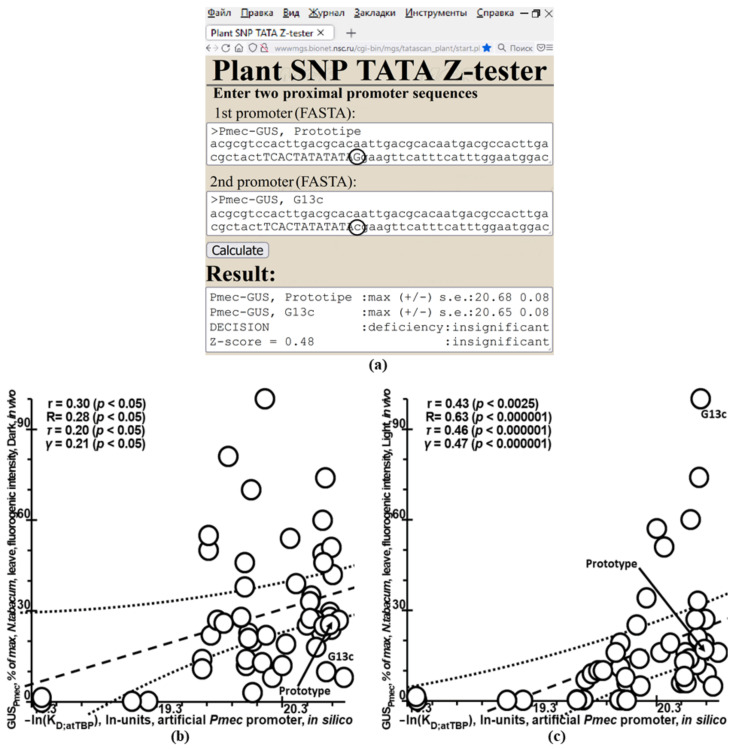
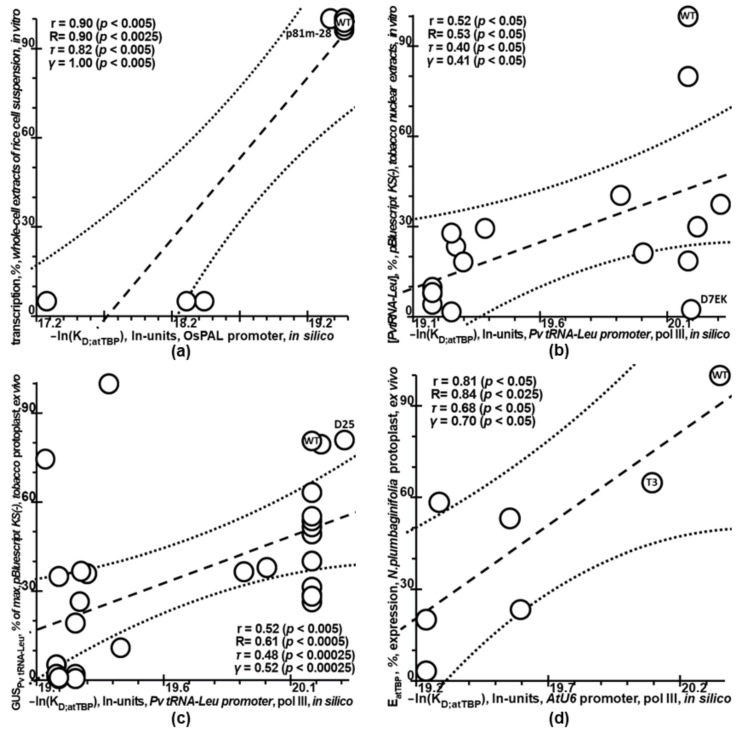
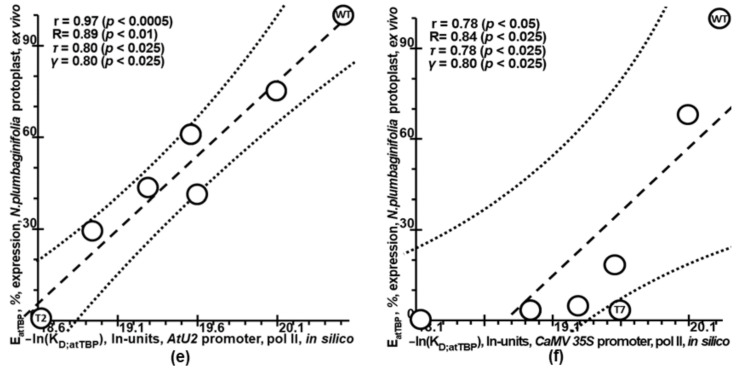
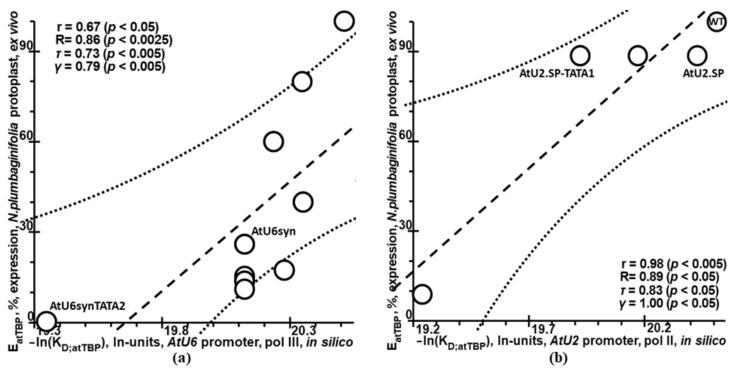
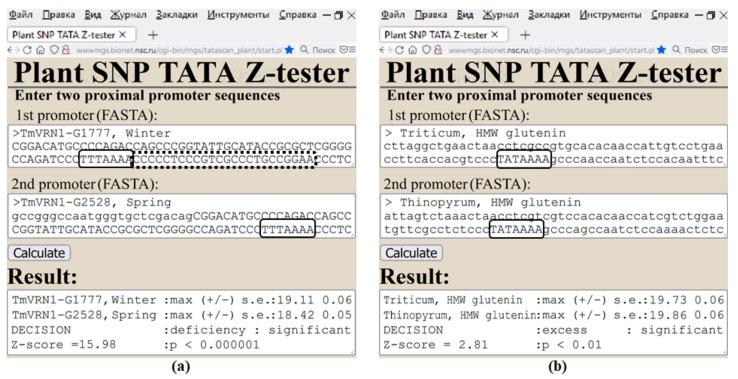
Synthetic targeted optimization of plant promoters is becoming a part of progress in mainstream postgenomic agriculture along with hybridization of cultivated plants with wild congeners, as well as marker-assisted breeding. Therefore, here, for the first time, we compiled all the experimental data-on mutational effects in plant proximal promoters on gene expression-that we could find in PubMed. Some of these datasets cast doubt on both the existence and the uniqueness of the sought solution, which could unequivocally estimate effects of proximal promoter mutation on gene expression when plants are grown under various environmental conditions during their development. This means that the inverse problem under study is ill-posed. Furthermore, we found experimental data on in vitro interchangeability of plant and human TATA-binding proteins allowing the application of Tikhonov's regularization, making this problem well-posed. Within these frameworks, we created our Web service Plant_SNP_TATA_Z-tester and then determined the limits of its applicability using those data that cast doubt on both the existence and the uniqueness of the sought solution. We confirmed that the effects (of proximal promoter mutations on gene expression) predicted by Plant_SNP_TATA_Z-tester correlate statistically significantly with all the experimental data under study. Lastly, we exemplified an application of Plant_SNP_TATA_Z-tester to agriculturally valuable mutations in plant promoters.
Keywords: TATA box; TATA-binding protein; Web service; correlation; development; environmental exposure; expression; gene; marker-assisted breeding; mutation; plant; plant hybrid; prediction; promoter; verification.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
AtSNP_TATAdb: Candidate Molecular Markers of Plant Advantages Related to Single Nucleotide Polymorphisms within Proximal Promoters of Arabidopsis thaliana L.Int J Mol Sci. 2024 Jan 3;25(1):607. doi: 10.3390/ijms25010607. Int J Mol Sci. 2024. PMID: 38203780 Free PMC article.
-
Promoters of genes encoding β-amylase, albumin and globulin in food plants have weaker affinity for TATA-binding protein as compared to non-food plants: in silico analysis.Vavilovskii Zhurnal Genet Selektsii. 2022 Dec;26(8):798-805. doi: 10.18699/VJGB-22-96. Vavilovskii Zhurnal Genet Selektsii. 2022. PMID: 36694715 Free PMC article.
-
Disruptive natural selection by male reproductive potential prevents underexpression of protein-coding genes on the human Y chromosome as a self-domestication syndrome.BMC Genet. 2020 Oct 22;21(Suppl 1):89. doi: 10.1186/s12863-020-00896-6. BMC Genet. 2020. PMID: 33092533 Free PMC article.
-
Human_SNP_TATAdb: a database of SNPs that statistically significantly change the affinity of the TATA-binding protein to human gene promoters: genome-wide analysis and use cases.Vavilovskii Zhurnal Genet Selektsii. 2023 Dec;27(7):728-736. doi: 10.18699/VJGB-23-85. Vavilovskii Zhurnal Genet Selektsii. 2023. PMID: 38213714 Free PMC article.
-
TATA box polymorphisms in human gene promoters and associated hereditary pathologies.Biochemistry (Mosc). 2009 Feb;74(2):117-29. doi: 10.1134/s0006297909020011. Biochemistry (Mosc). 2009. PMID: 19267666 Review.
Cited by
-
AtSNP_TATAdb: Candidate Molecular Markers of Plant Advantages Related to Single Nucleotide Polymorphisms within Proximal Promoters of Arabidopsis thaliana L.Int J Mol Sci. 2024 Jan 3;25(1):607. doi: 10.3390/ijms25010607. Int J Mol Sci. 2024. PMID: 38203780 Free PMC article.
-
Candidate SNP Markers Significantly Altering the Affinity of TATA-Binding Protein for the Promoters of Human Hub Genes for Atherogenesis, Atherosclerosis and Atheroprotection.Int J Mol Sci. 2023 May 19;24(10):9010. doi: 10.3390/ijms24109010. Int J Mol Sci. 2023. PMID: 37240358 Free PMC article.
-
Developmental Biology: Computational and Experimental Approaches.Int J Mol Sci. 2023 Jun 21;24(13):10435. doi: 10.3390/ijms241310435. Int J Mol Sci. 2023. PMID: 37445614 Free PMC article.
-
Research Topics of the Bioinformatics of Gene Regulation.Int J Mol Sci. 2023 May 15;24(10):8774. doi: 10.3390/ijms24108774. Int J Mol Sci. 2023. PMID: 37240120 Free PMC article.
-
Differentially Expressed Genes and Molecular Susceptibility to Human Age-Related Diseases.Int J Mol Sci. 2023 Feb 16;24(4):3996. doi: 10.3390/ijms24043996. Int J Mol Sci. 2023. PMID: 36835409 Free PMC article.
References
-
- Srivastava R.K., Yadav O.P., Kaliamoorthy S., Gupta S.K., Serba D.D., Choudhary S., Govindaraj M., Kholova J., Murugesan T., Satyavathi C.T., et al. Breeding drought-tolerant pearl millet using conventional and genomic approaches: Achievements and prospects. Front. Plant Sci. 2022;13:781524. doi: 10.3389/fpls.2022.781524. - DOI - PMC - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
