

The Rise and Fall of Billionaire siRNAs during Reproductive Development in Rice
- PMID: 35956435
- PMCID: PMC9370656
- DOI: 10.3390/plants11151957
The Rise and Fall of Billionaire siRNAs during Reproductive Development in Rice
Abstract
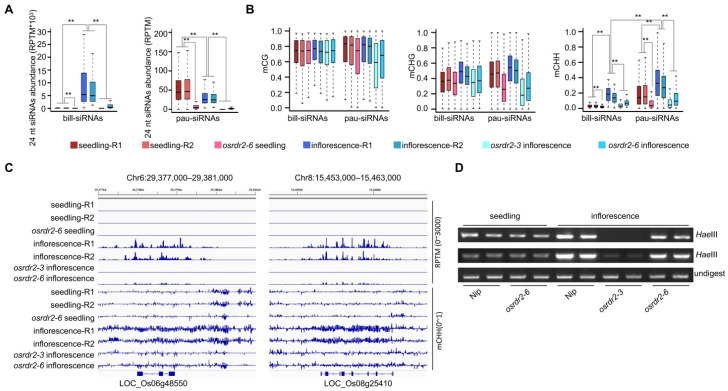
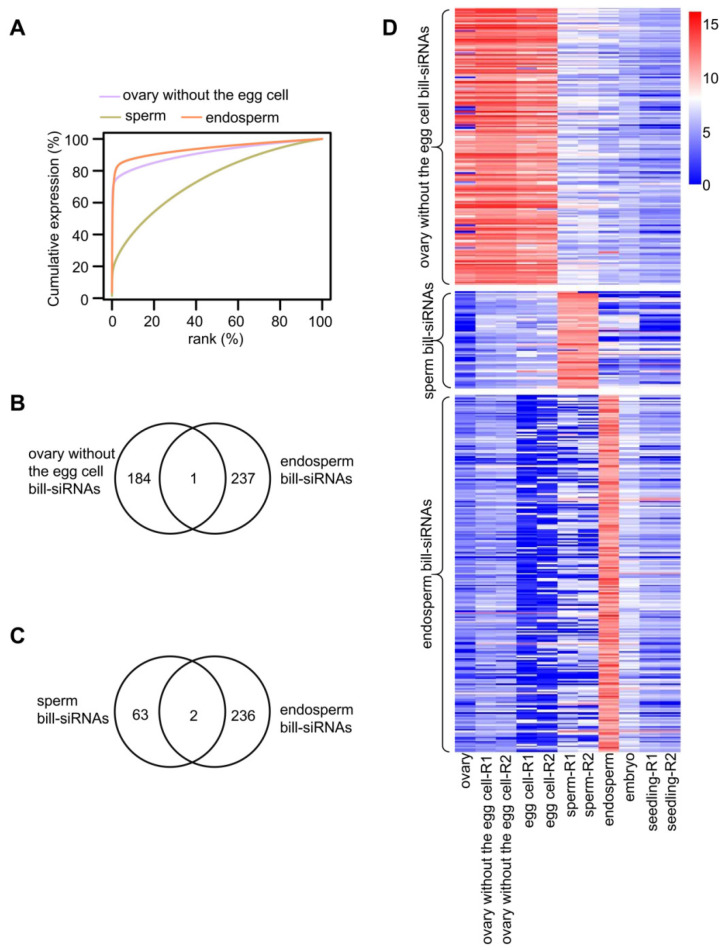
RNA polymerase IV-dependent siRNAs, usually 24 nt in length, function in the RNA-directed DNA methylation that is responsible for de novo methylation in plants. We analyzed 24 nt siRNAs in inflorescences and found that among the 20,200 24 nt siRNA clusters, the top 0.81% highly expressed clusters accounted for more than 68% of the 24 nt siRNA reads in inflorescences. We named the highly expressed siRNAs as billionaire siRNAs (bill-siRNAs) and the less-expressed siRNAs as pauper siRNAs (pau-siRNAs). The bill-siRNAs in inflorescences are mainly derived from the ovary. Female gametes produced more bill-siRNAs than male gametes. In embryos and seedlings developed from fertilized egg cells, the bill-siRNAs from gametes disappeared. The endosperm, which develops from the fertilized central cell, also contained no bill-siRNAs from gametes but did contain newly and highly expressed siRNAs produced in different regions. In contrast, bill-siRNAs from the ovaries were maintained in the seed coat. The biosynthesis of bill-siRNAs in various tissues and cells is dependent on OsRDR2 (RNA-dependent RNA polymerase 2) and Pol IV (DNA-dependent RNA polymerase IV). Similar to the pau-siRNAs, the first base of bill-siRNAs is enriched at adenine, and bill-siRNAs can direct DNA methylation in various tissues.
Keywords: reproductive development; rice; siRNA.
Conflict of interest statement
The authors declare no conflict of interest.
Figures






Similar articles
-
The effect of RNA polymerase V on 24-nt siRNA accumulation depends on DNA methylation contexts and histone modifications in rice.Proc Natl Acad Sci U S A. 2021 Jul 27;118(30):e2100709118. doi: 10.1073/pnas.2100709118. Proc Natl Acad Sci U S A. 2021. PMID: 34290143 Free PMC article.
-
Distinct and concurrent pathways of Pol II- and Pol IV-dependent siRNA biogenesis at a repetitive trans-silencer locus in Arabidopsis thaliana.Plant J. 2014 Jul;79(1):127-38. doi: 10.1111/tpj.12545. Epub 2014 Jun 13. Plant J. 2014. PMID: 24798377
-
Abundant expression of maternal siRNAs is a conserved feature of seed development.Proc Natl Acad Sci U S A. 2020 Jun 30;117(26):15305-15315. doi: 10.1073/pnas.2001332117. Epub 2020 Jun 15. Proc Natl Acad Sci U S A. 2020. PMID: 32541052 Free PMC article.
-
RNA-directed DNA methylation in plants: Where to start?RNA Biol. 2013 Oct;10(10):1593-6. doi: 10.4161/rna.26312. RNA Biol. 2013. PMID: 25003825 Free PMC article. Review.
-
Advances of RNA polymerase IV in controlling DNA methylation and development in plants.Yi Chuan. 2022 Jul 20;44(7):567-580. doi: 10.16288/j.yczz.22-063. Yi Chuan. 2022. PMID: 35858769 Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
