

Dynamics of Host Immune Response Development During Schistosoma mansoni Infection
- PMID: 35958580
- PMCID: PMC9362740
- DOI: 10.3389/fimmu.2022.906338
Dynamics of Host Immune Response Development During Schistosoma mansoni Infection
Erratum in
-
Corrigendum: Dynamics of host immune response development during Schistosoma mansoni infection.Front Immunol. 2023 Aug 10;14:1229665. doi: 10.3389/fimmu.2023.1229665. eCollection 2023. Front Immunol. 2023. PMID: 37675098 Free PMC article.
Abstract
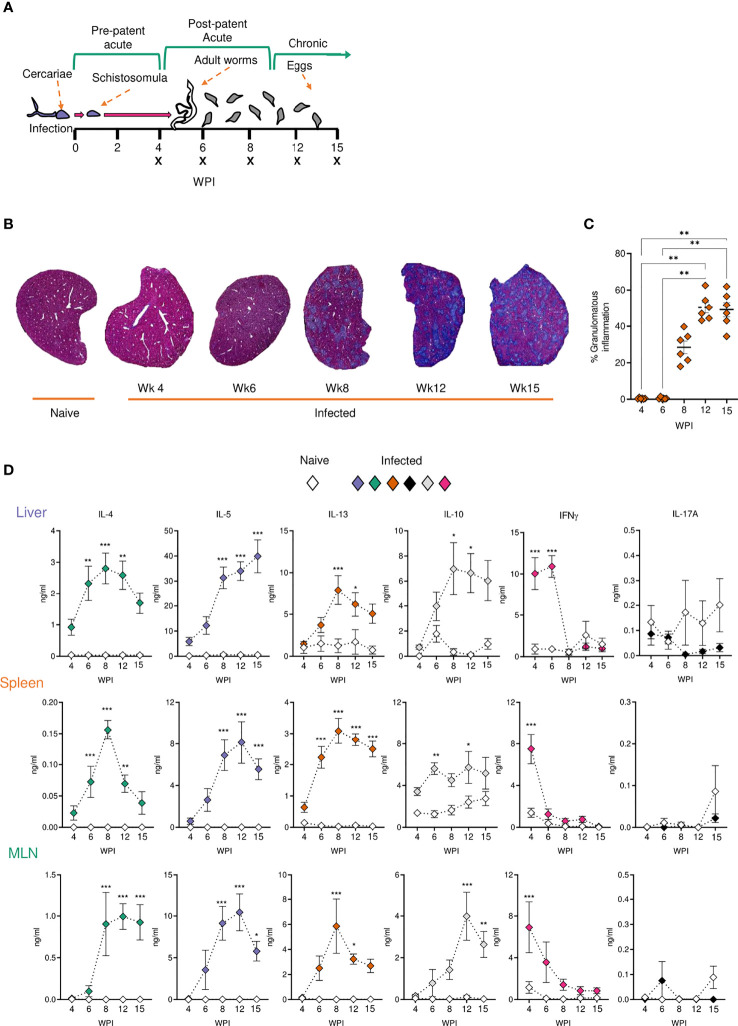
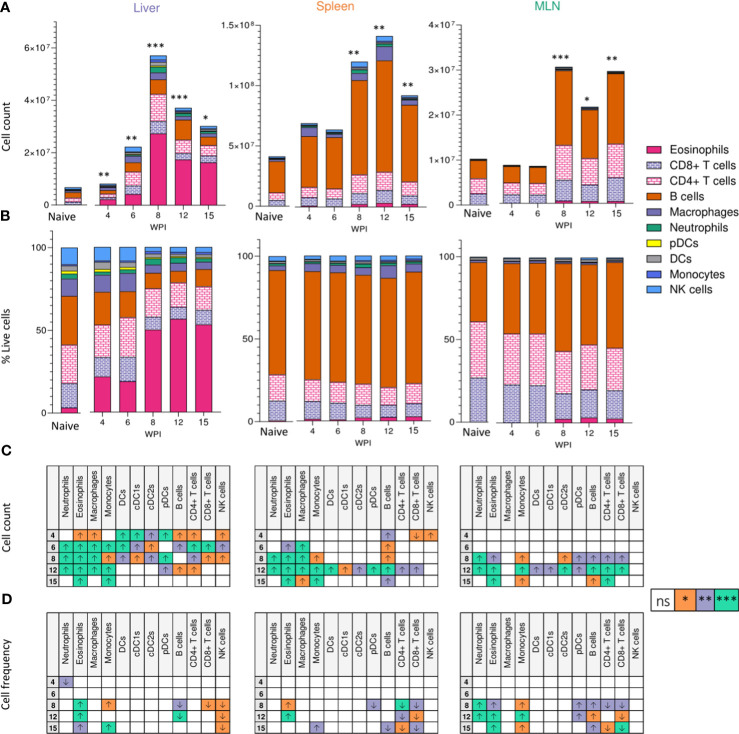
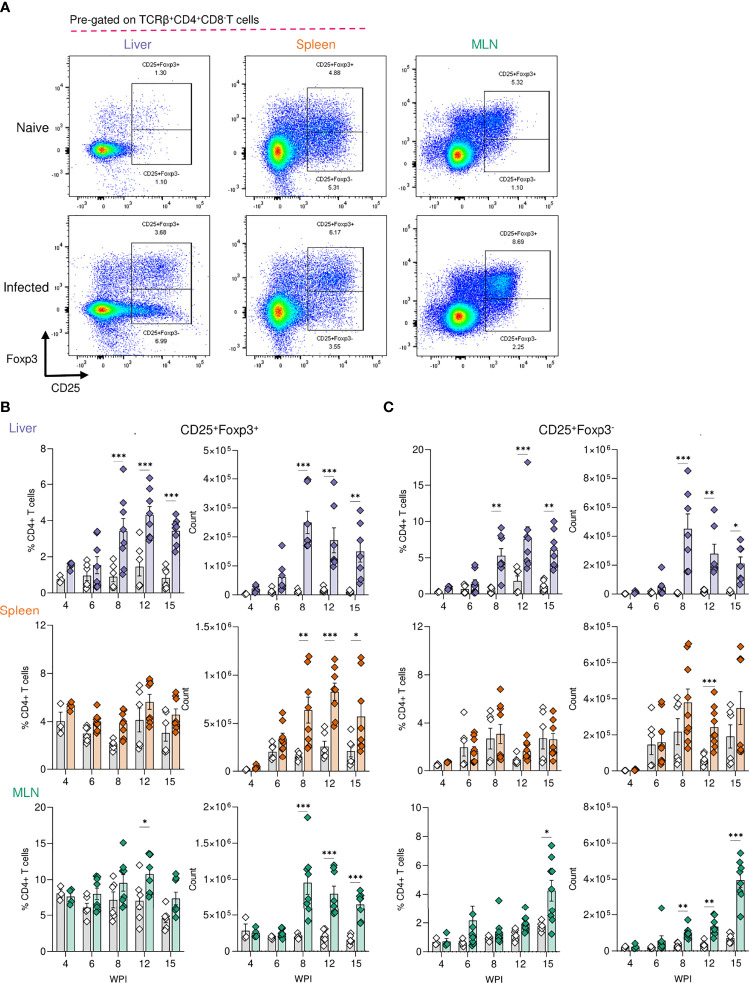
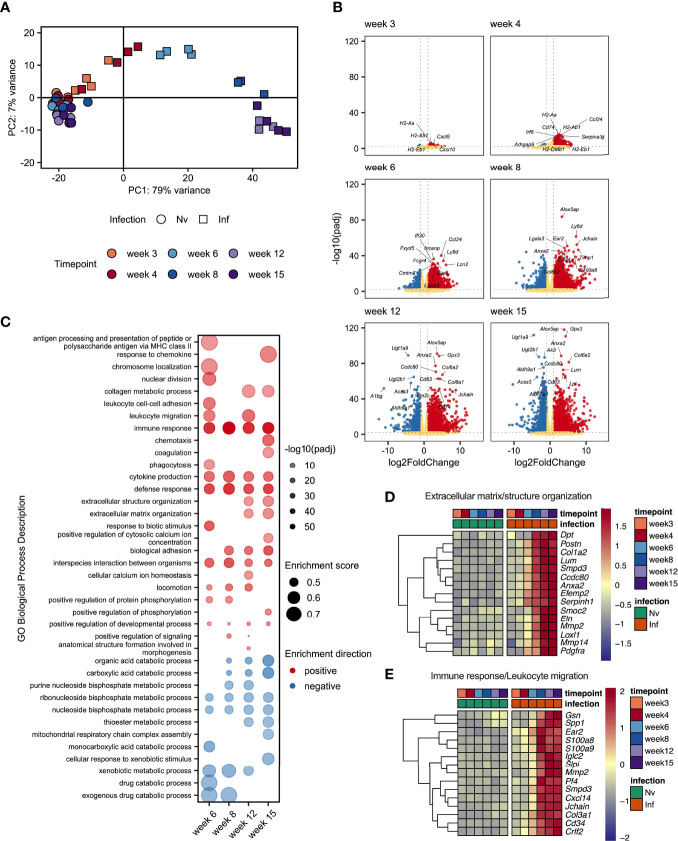
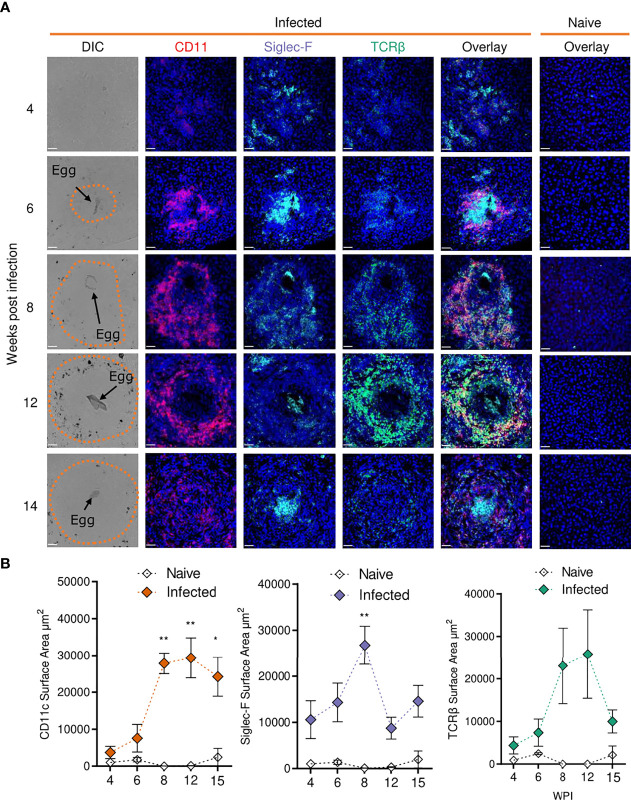
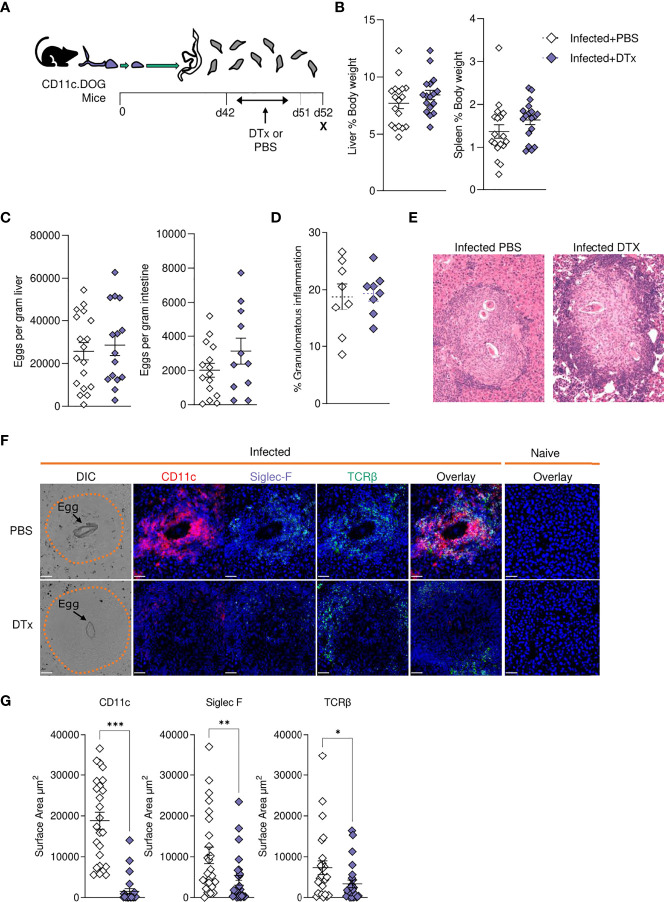
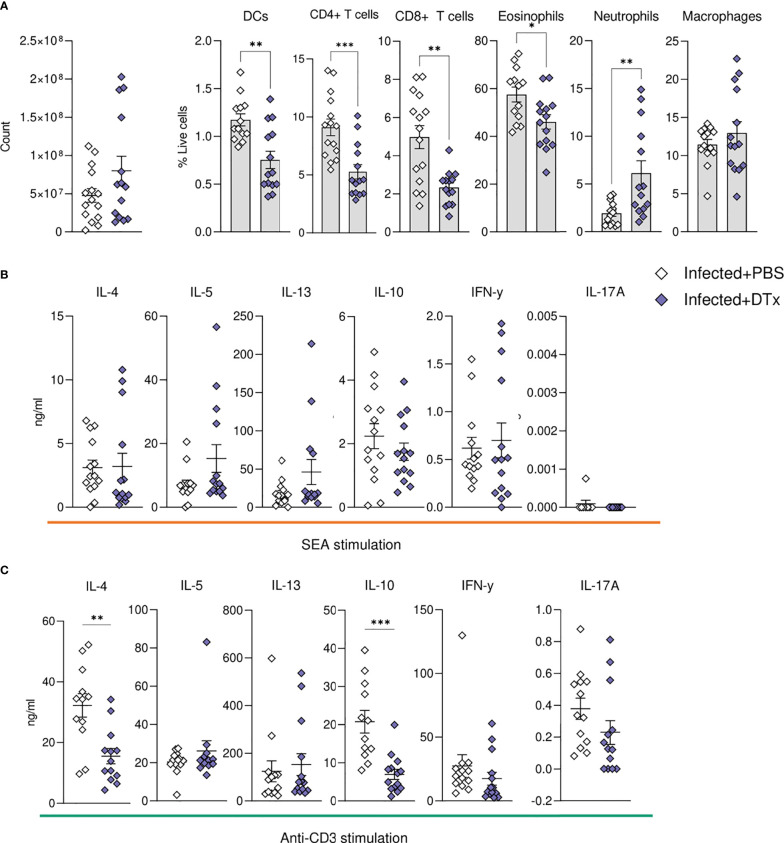
Schistosomiasis is a disease of global significance, with severity and pathology directly related to how the host responds to infection. The immunological narrative of schistosomiasis has been constructed through decades of study, with researchers often focussing on isolated time points, cell types and tissue sites of interest. However, the field currently lacks a comprehensive and up-to-date understanding of the immune trajectory of schistosomiasis over infection and across multiple tissue sites. We have defined schistosome-elicited immune responses at several distinct stages of the parasite lifecycle, in three tissue sites affected by infection: the liver, spleen, and mesenteric lymph nodes. Additionally, by performing RNA-seq on the livers of schistosome infected mice, we have generated novel transcriptomic insight into the development of schistosome-associated liver pathology and fibrosis across the breadth of infection. Through depletion of CD11c+ cells during peak stages of schistosome-driven inflammation, we have revealed a critical role for CD11c+ cells in the co-ordination and regulation of Th2 inflammation during infection. Our data provide an updated and high-resolution account of how host immune responses evolve over the course of murine schistosomiasis, underscoring the significance of CD11c+ cells in dictating host immunopathology against this important helminth infection.
Keywords: chronic infection; dendritic cells; pathology; schistosomiasis; transcriptomic (RNA-seq).
Copyright © 2022 Costain, Phythian-Adams, Colombo, Marley, Owusu, Cook, Brown, Webb, Lundie, Smits, Berriman and MacDonald.
Conflict of interest statement
Author RL was employed by company 360biolabs. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
Defined Intestinal Regions Are Drained by Specific Lymph Nodes That Mount Distinct Th1 and Th2 Responses Against Schistosoma mansoni Eggs.Front Immunol. 2020 Oct 23;11:592325. doi: 10.3389/fimmu.2020.592325. eCollection 2020. Front Immunol. 2020. PMID: 33193437 Free PMC article.
-
Strongyloides venezuelensis-infection alters the profile of cytokines and liver inflammation in mice co-infected with Schistosoma mansoni.Cytokine. 2020 Mar;127:154931. doi: 10.1016/j.cyto.2019.154931. Epub 2019 Nov 26. Cytokine. 2020. PMID: 31783260
-
IL-4Rα-expressing CD11c+ cells contribute to driving optimal cellular responses during Schistosoma mansoni infection in mice.J Leukoc Biol. 2019 Feb;105(2):307-316. doi: 10.1002/JLB.MA0318-115R. Epub 2018 Nov 30. J Leukoc Biol. 2019. PMID: 30500088 Free PMC article.
-
The C-type Lectin Receptor-Driven, Th17 Cell-Mediated Severe Pathology in Schistosomiasis: Not All Immune Responses to Helminth Parasites Are Th2 Dominated.Front Immunol. 2019 Jan 30;10:26. doi: 10.3389/fimmu.2019.00026. eCollection 2019. Front Immunol. 2019. PMID: 30761125 Free PMC article. Review.
-
Cellular and chemokine-mediated regulation in schistosome-induced hepatic pathology.Trends Parasitol. 2014 Mar;30(3):141-50. doi: 10.1016/j.pt.2013.12.009. Epub 2014 Jan 13. Trends Parasitol. 2014. PMID: 24433721 Review.
Cited by
-
Clinical tolerance but no protective efficacy in a placebo-controlled trial of repeated controlled schistosome infection.J Clin Invest. 2024 Dec 12;135(4):e185422. doi: 10.1172/JCI185422. J Clin Invest. 2024. PMID: 39666392 Free PMC article. Clinical Trial.
-
Evaluation of IL-35, as a Possible Biomarker for Follow-Up after Therapy, in Chronic Human Schistosoma Infection.Vaccines (Basel). 2023 May 17;11(5):995. doi: 10.3390/vaccines11050995. Vaccines (Basel). 2023. PMID: 37243099 Free PMC article.
-
Pre-clinical studies of Schistosoma mansoni vaccines: A scoping review.PLoS Negl Trop Dis. 2025 Jun 2;19(6):e0012956. doi: 10.1371/journal.pntd.0012956. eCollection 2025 Jun. PLoS Negl Trop Dis. 2025. PMID: 40455825 Free PMC article.
-
Schistosomiasis Mansoni-Recruited Eosinophils: An Overview in the Granuloma Context.Microorganisms. 2022 Oct 13;10(10):2022. doi: 10.3390/microorganisms10102022. Microorganisms. 2022. PMID: 36296298 Free PMC article. Review.
-
Regulation of immune response against third-stage Gnathostoma spinigerum larvae by human genes.Front Immunol. 2023 Aug 3;14:1218965. doi: 10.3389/fimmu.2023.1218965. eCollection 2023. Front Immunol. 2023. PMID: 37600806 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials