

The matricellular protein SPARC induces inflammatory interferon-response in macrophages during aging
- PMID: 35963236
- PMCID: PMC9474643
- DOI: 10.1016/j.immuni.2022.07.007
The matricellular protein SPARC induces inflammatory interferon-response in macrophages during aging
Abstract
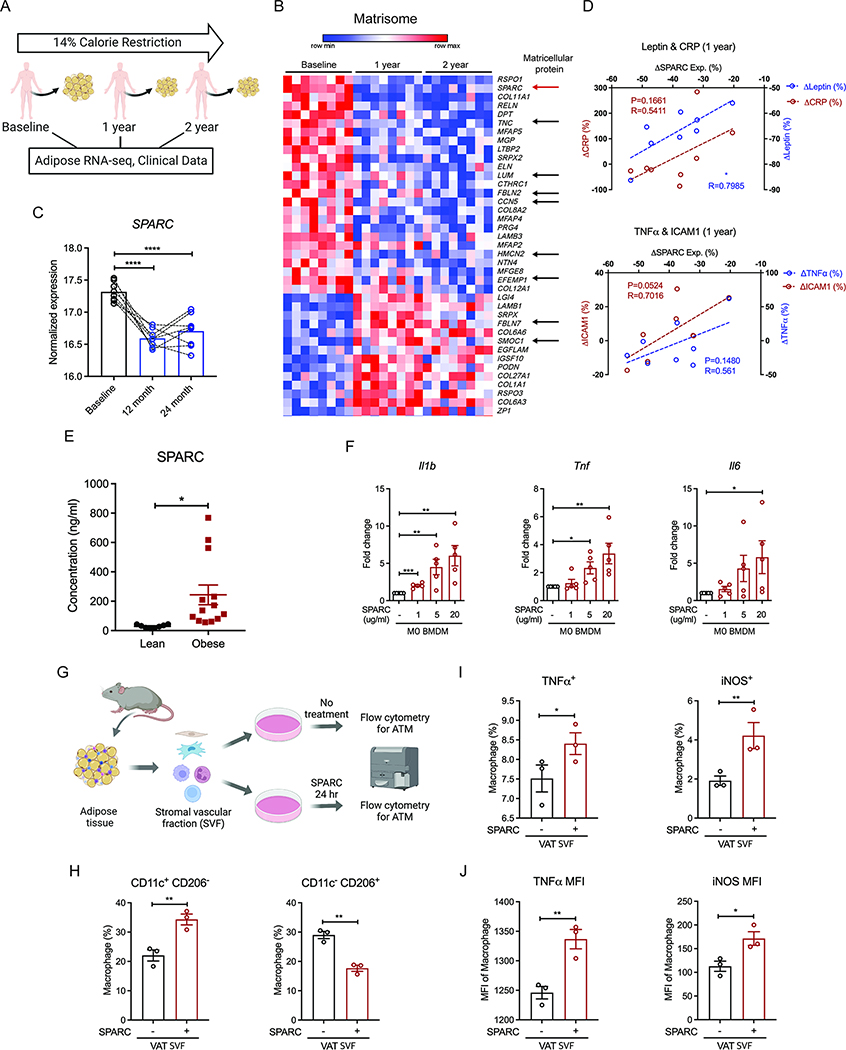
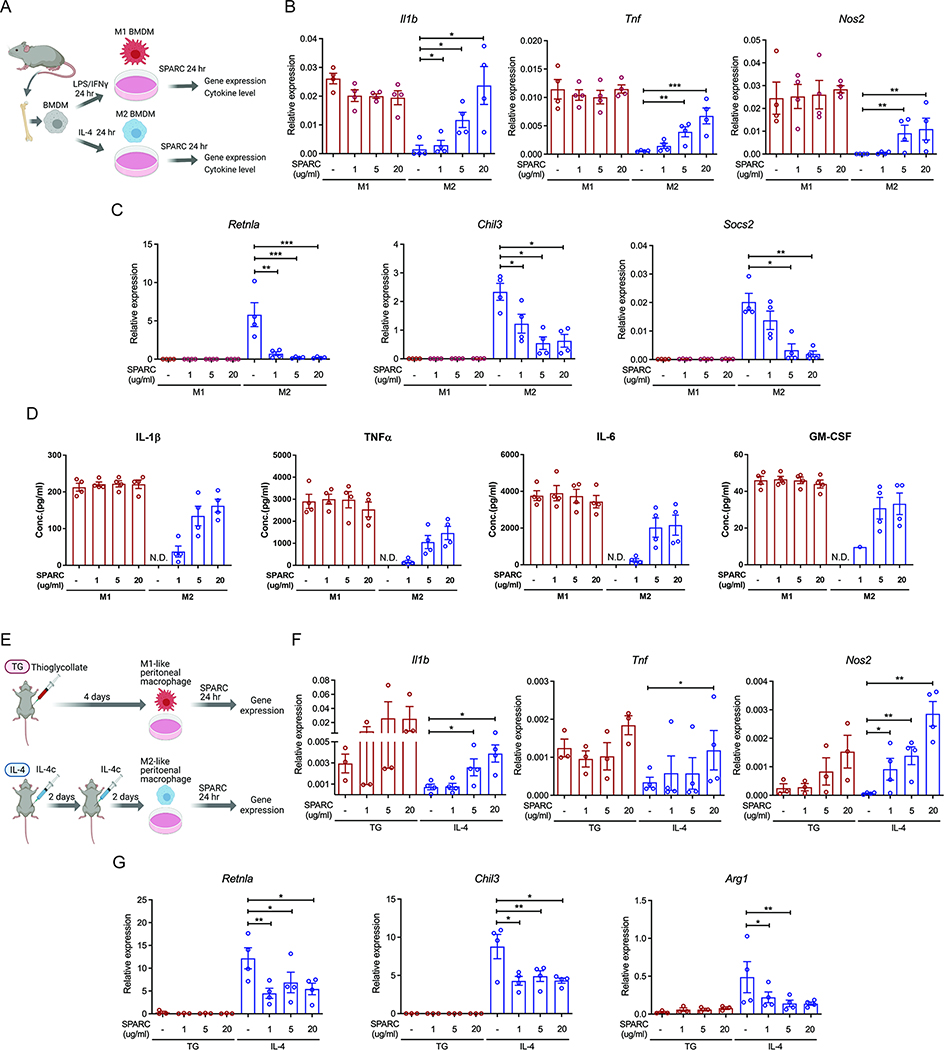
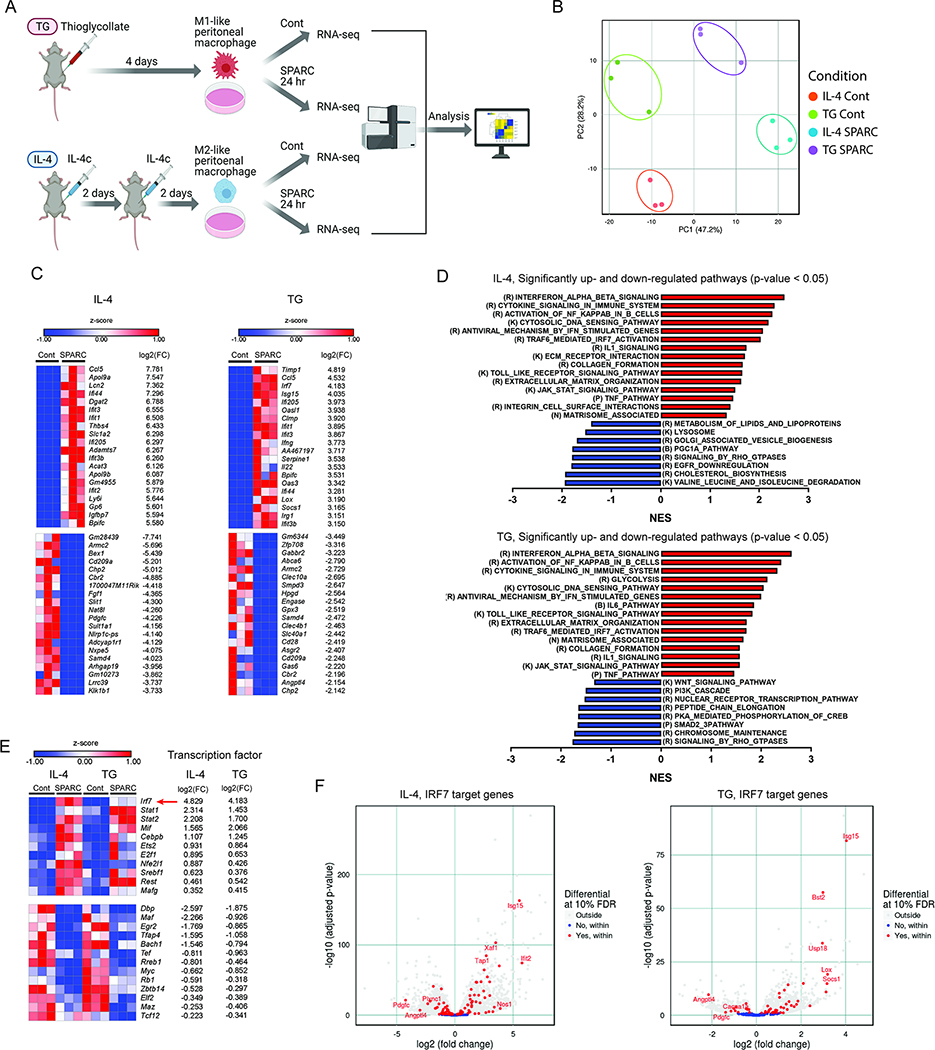
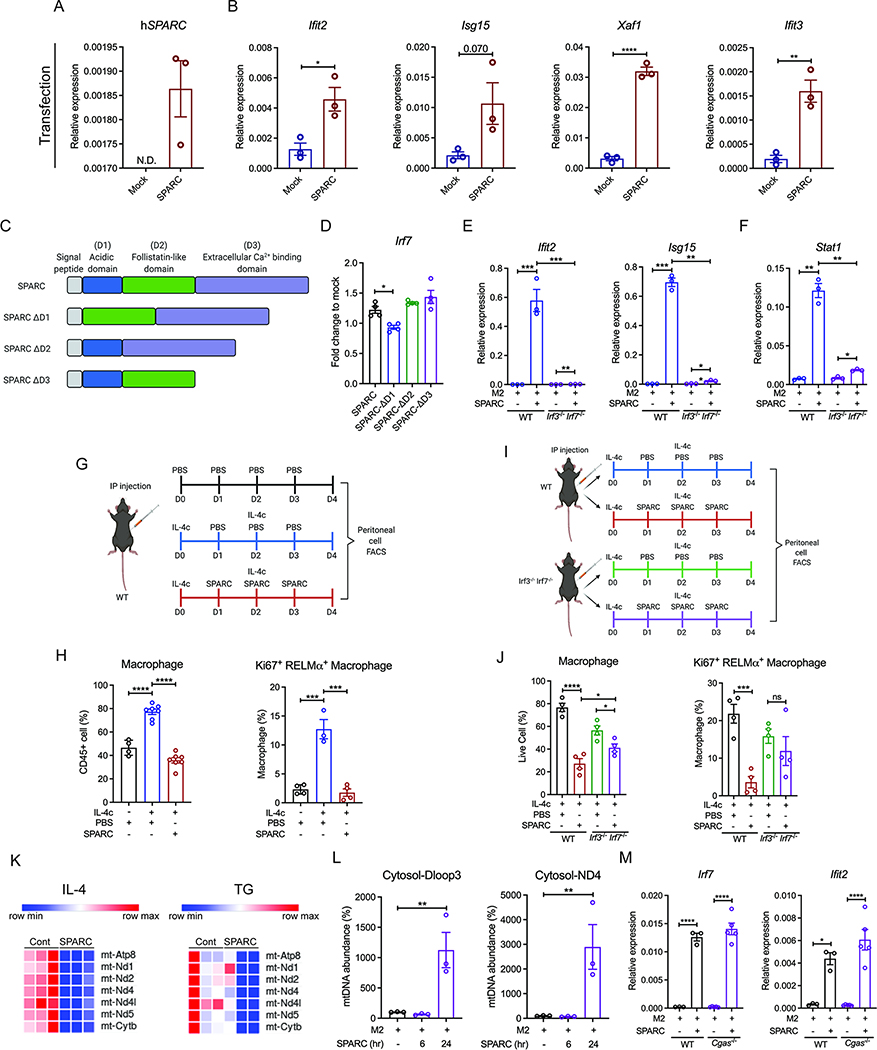
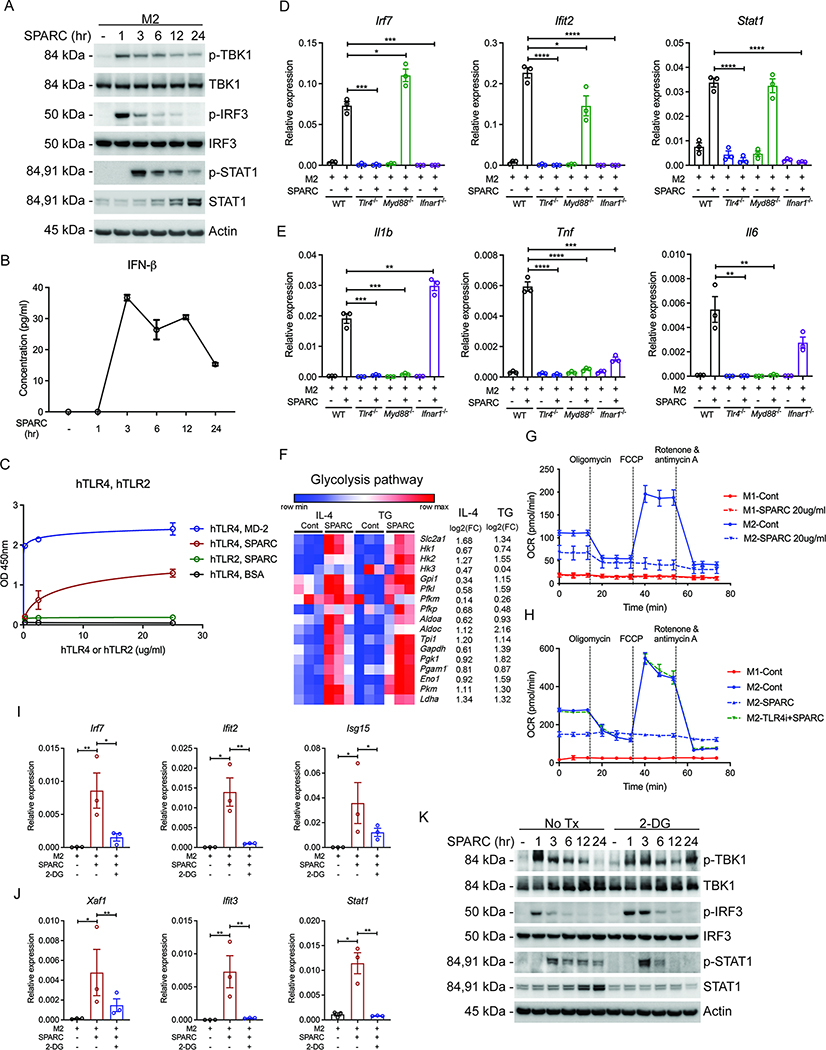
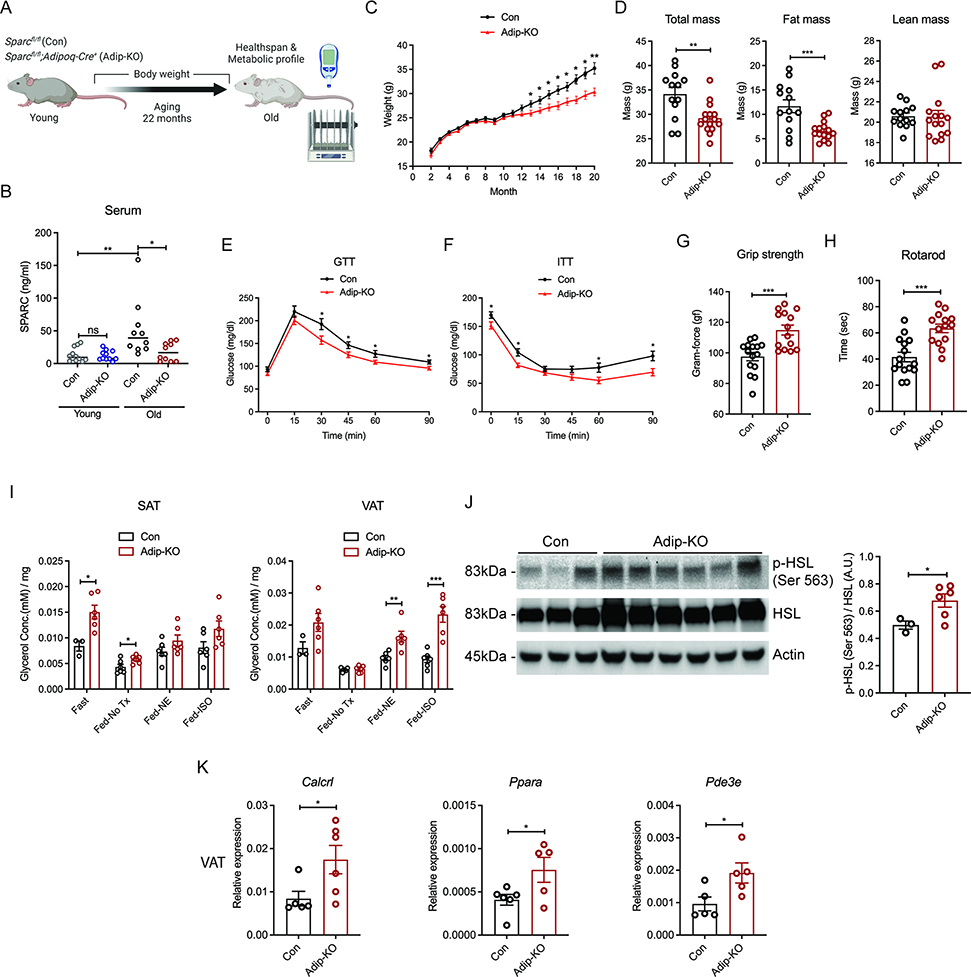
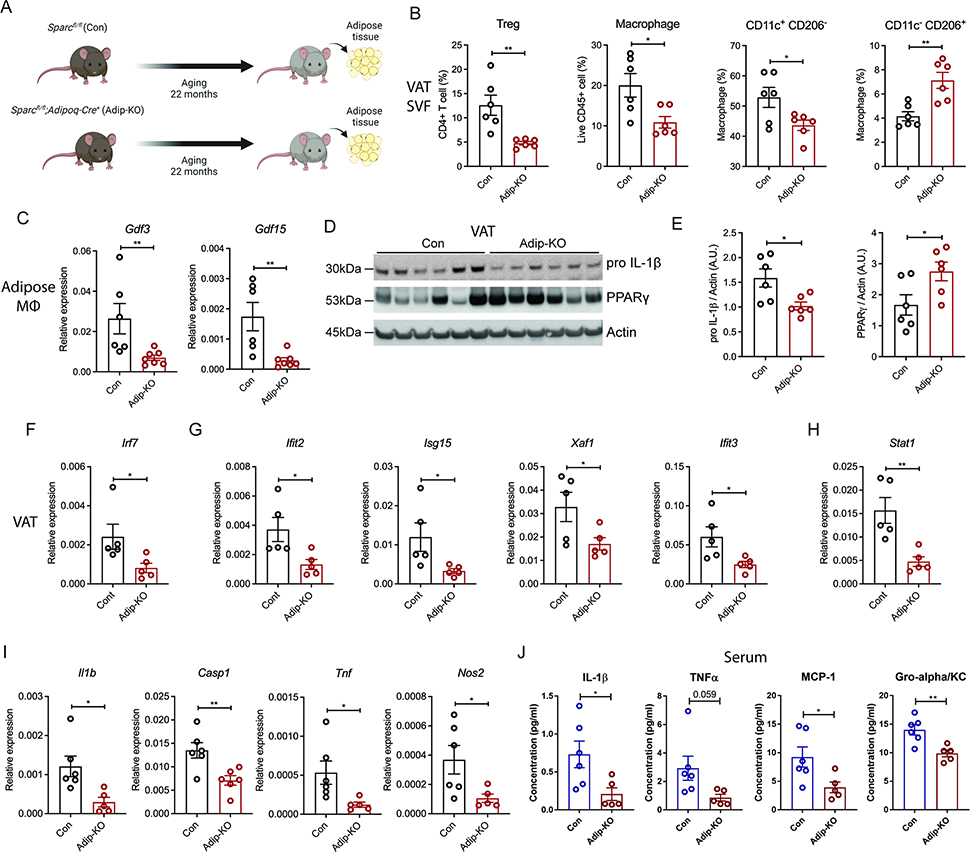
The risk of chronic diseases caused by aging is reduced by caloric restriction (CR)-induced immunometabolic adaptation. Here, we found that the matricellular protein, secreted protein acidic and rich in cysteine (SPARC), was inhibited by 2 years of 14% sustained CR in humans and elevated by obesity. SPARC converted anti-inflammatory macrophages into a pro-inflammatory phenotype with induction of interferon-stimulated gene (ISG) expression via the transcription factors IRF3/7. Mechanistically, SPARC-induced ISGs were dependent on toll-like receptor-4 (TLR4)-mediated TBK1, IRF3, IFN-β, and STAT1 signaling without engaging the Myd88 pathway. Metabolically, SPARC dampened mitochondrial respiration, and inhibition of glycolysis abrogated ISG induction by SPARC in macrophages. Furthermore, the N-terminal acidic domain of SPARC was required for ISG induction, while adipocyte-specific deletion of SPARC reduced inflammation and extended health span during aging. Collectively, SPARC, a CR-mimetic adipokine, is an immunometabolic checkpoint of inflammation and interferon response that may be targeted to delay age-related metabolic and functional decline.
Keywords: SPARC; TLR4; caloric restriction; inflammation; interferon-stimulated gene; macrophage; matricellular protein.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Skip the buffet, for SPARC's sake.Immunity. 2022 Sep 13;55(9):1583-1585. doi: 10.1016/j.immuni.2022.08.012. Immunity. 2022. PMID: 36103855
References
-
- Adler MI, Cassidy EJ, Fricke C, and Bonduriansky R (2013). The lifespan-reproduction trade-off under dietary restriction is sex-specific and context-dependent. Exp Gerontol 48, 539–548. - PubMed
-
- Bai J, Cervantes C, Liu J, He S, Zhou H, Zhang B, Cai H, Yin D, Hu D, Li Z, et al. (2017). DsbA-L prevents obesity-induced inflammation and insulin resistance by suppressing the mtDNA release-activated cGAS-cGAMP-STING pathway. Proceedings of the National Academy of Sciences 114, 12196–12201. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 AR070811/AR/NIAMS NIH HHS/United States
- U01 AG020478/AG/NIA NIH HHS/United States
- U01 AG022132/AG/NIA NIH HHS/United States
- P30 DK072476/DK/NIDDK NIH HHS/United States
- U54 GM104940/GM/NIGMS NIH HHS/United States
- P01 AG051459/AG/NIA NIH HHS/United States
- R01 AG045712/AG/NIA NIH HHS/United States
- R01 AG068863/AG/NIA NIH HHS/United States
- R01 AG031797/AG/NIA NIH HHS/United States
- R01 AG073969/AG/NIA NIH HHS/United States
- U01 AG020487/AG/NIA NIH HHS/United States
- UL1 TR001863/TR/NCATS NIH HHS/United States
- U01 AG020480/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
