

Gliotransmission of D-serine promotes thirst-directed behaviors in Drosophila
- PMID: 35963239
- PMCID: PMC9616736
- DOI: 10.1016/j.cub.2022.07.038
Gliotransmission of D-serine promotes thirst-directed behaviors in Drosophila
Abstract
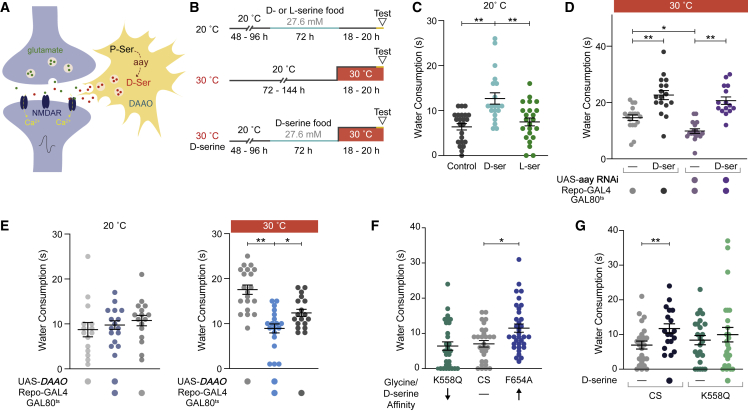
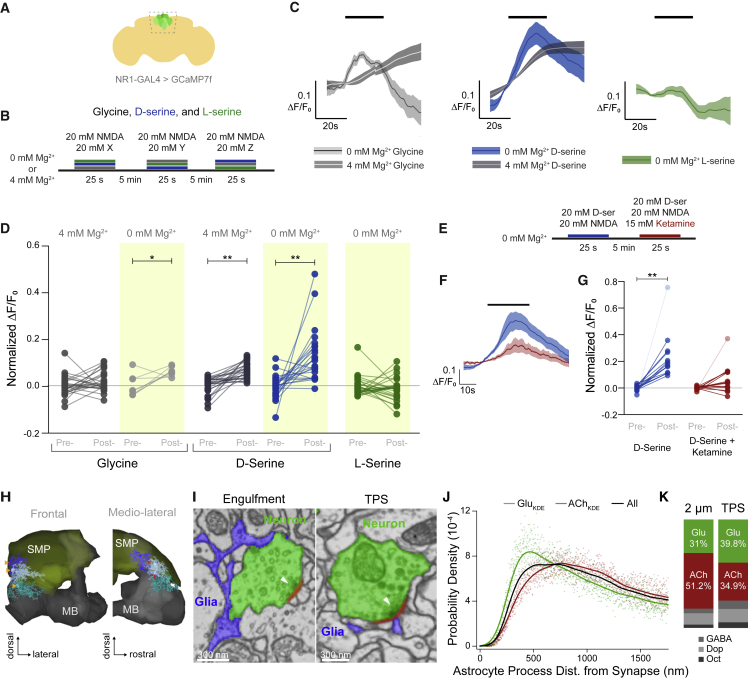
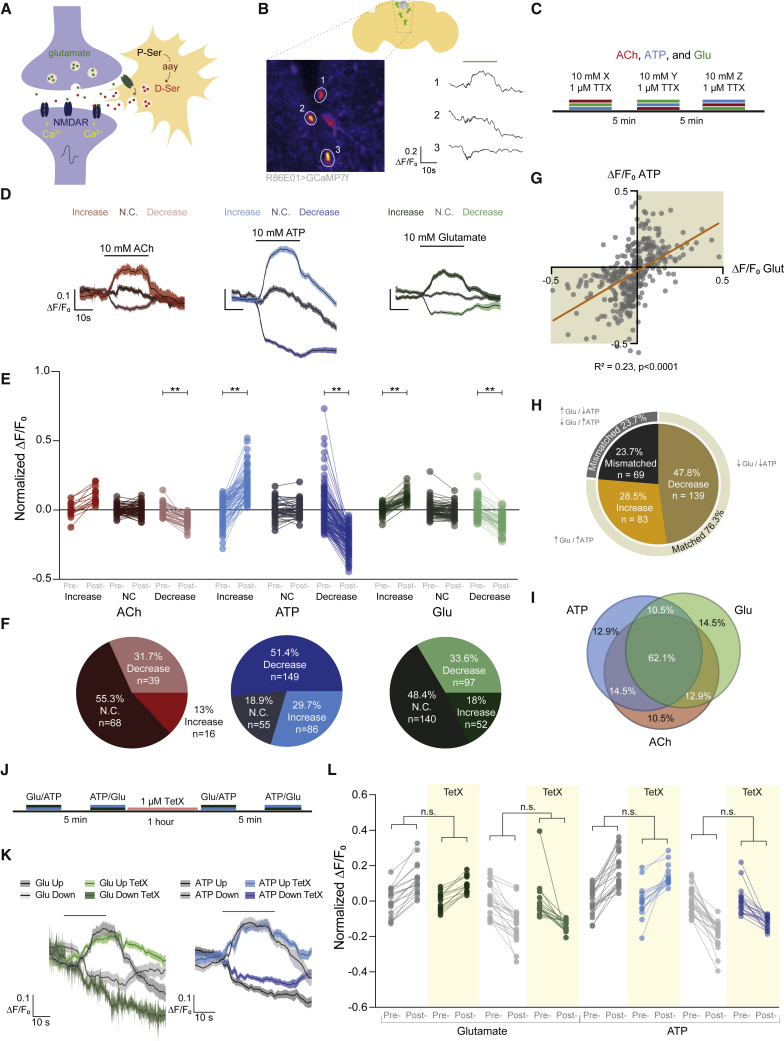
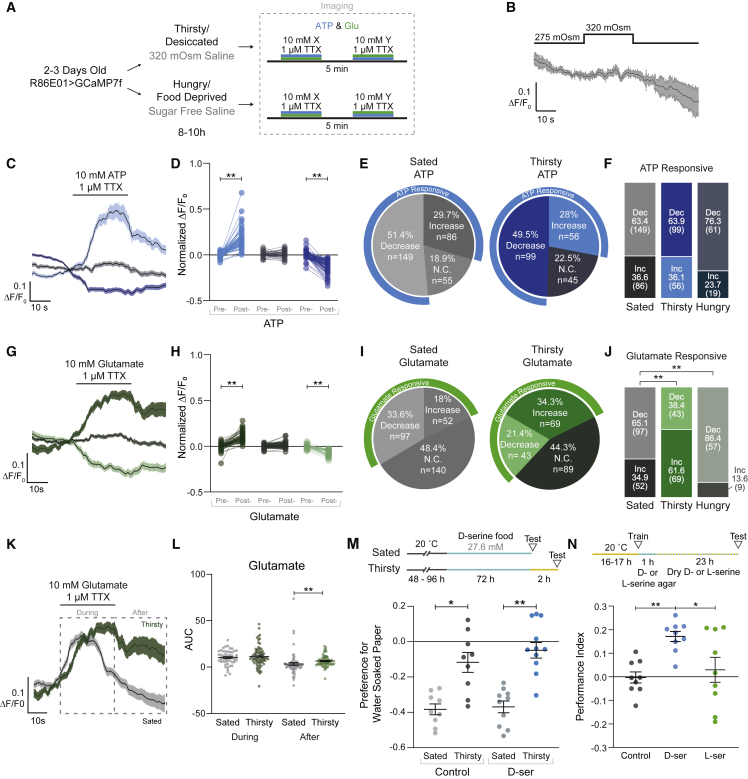
Thirst emerges from a range of cellular changes that ultimately motivate an animal to consume water. Although thirst-responsive neuronal signals have been reported, the full complement of brain responses is unclear. Here, we identify molecular and cellular adaptations in the brain using single-cell sequencing of water-deprived Drosophila. Water deficiency primarily altered the glial transcriptome. Screening the regulated genes revealed astrocytic expression of the astray-encoded phosphoserine phosphatase to bi-directionally regulate water consumption. Astray synthesizes the gliotransmitter D-serine, and vesicular release from astrocytes is required for drinking. Moreover, dietary D-serine rescues aay-dependent drinking deficits while facilitating water consumption and expression of water-seeking memory. D-serine action requires binding to neuronal NMDA-type glutamate receptors. Fly astrocytes contribute processes to tripartite synapses, and the proportion of astrocytes that are themselves activated by glutamate increases with water deprivation. We propose that thirst elevates astrocytic D-serine release, which awakens quiescent glutamatergic circuits to enhance water procurement.
Keywords: D-serine; Drosophila; NMDA receptors; astrocytes; behavior; glia; gliotransmission; single-cell transcriptomics; thirst; water procurement.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.W. is a member of the advisory board of Current Biology.
Figures
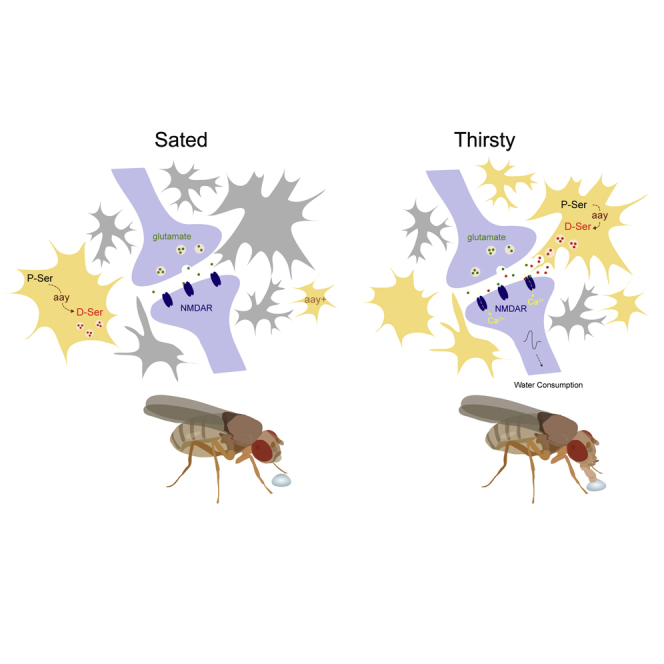
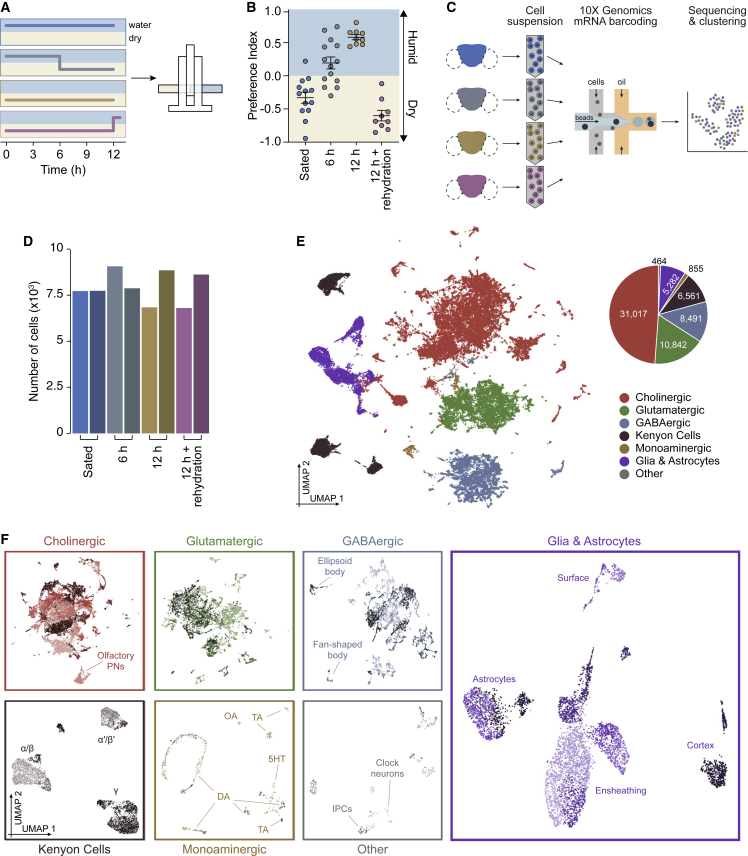
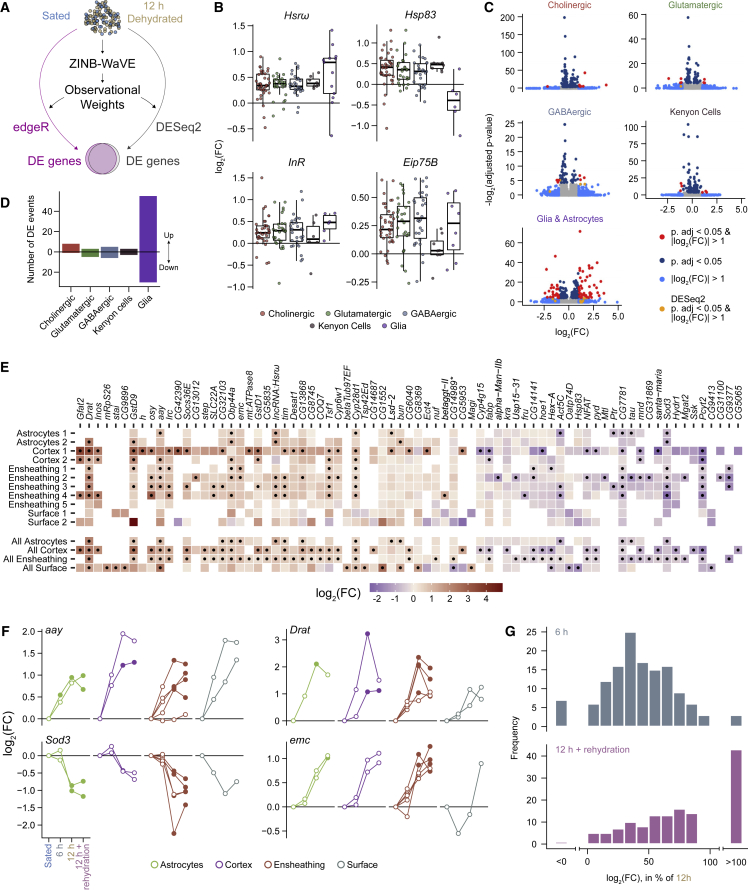
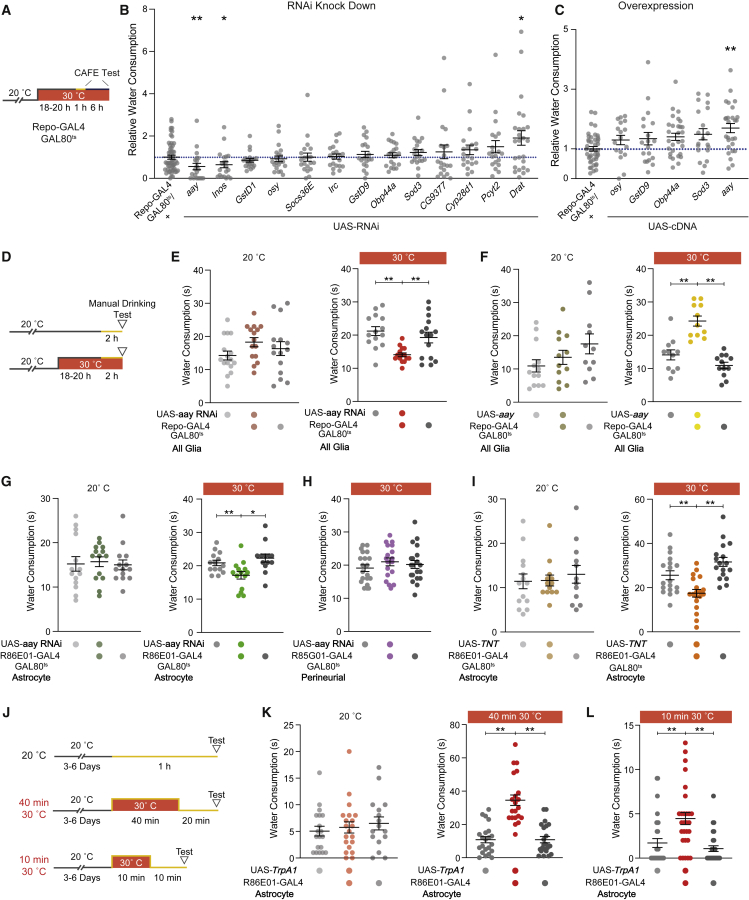
Comment in
-
Sensory biology: Thirsty glia motivate water consumption.Curr Biol. 2022 Sep 26;32(18):R949-R952. doi: 10.1016/j.cub.2022.08.048. Curr Biol. 2022. PMID: 36167042 Free PMC article.
References
-
- Bourque C.W. Central mechanisms of osmosensation and systemic osmoregulation. Nat. Rev. Neurosci. 2008;9:519–531. - PubMed
-
- Fitzsimons J.T. Angiotensin, thirst, and sodium appetite. Physiol. Rev. 1998;78:583–686. - PubMed
-
- Johnson A.K., Thunhorst R.L. The neuroendocrinology of thirst and salt appetite: visceral sensory signals and mechanisms of central integration. Front. Neuroendocrinol. 1997;18:292–353. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
