

SARS-CoV-2 spike N-terminal domain modulates TMPRSS2-dependent viral entry and fusogenicity
- PMID: 35963244
- PMCID: PMC9346021
- DOI: 10.1016/j.celrep.2022.111220
SARS-CoV-2 spike N-terminal domain modulates TMPRSS2-dependent viral entry and fusogenicity
Abstract
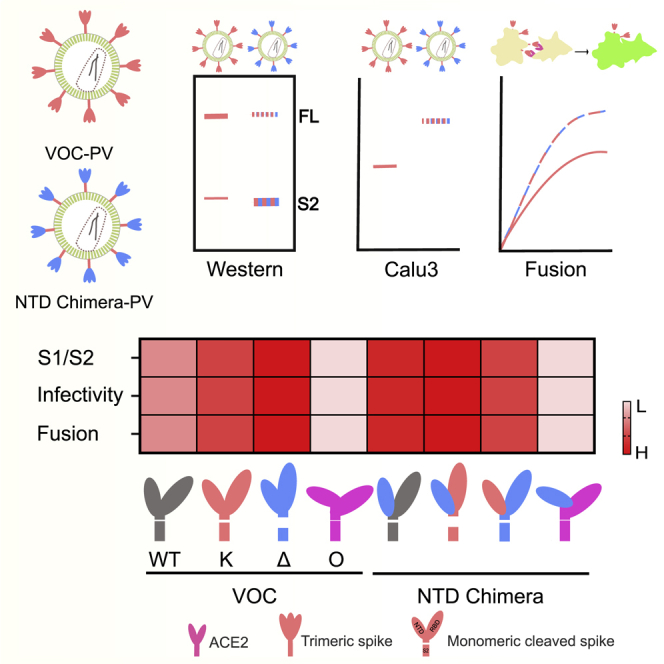
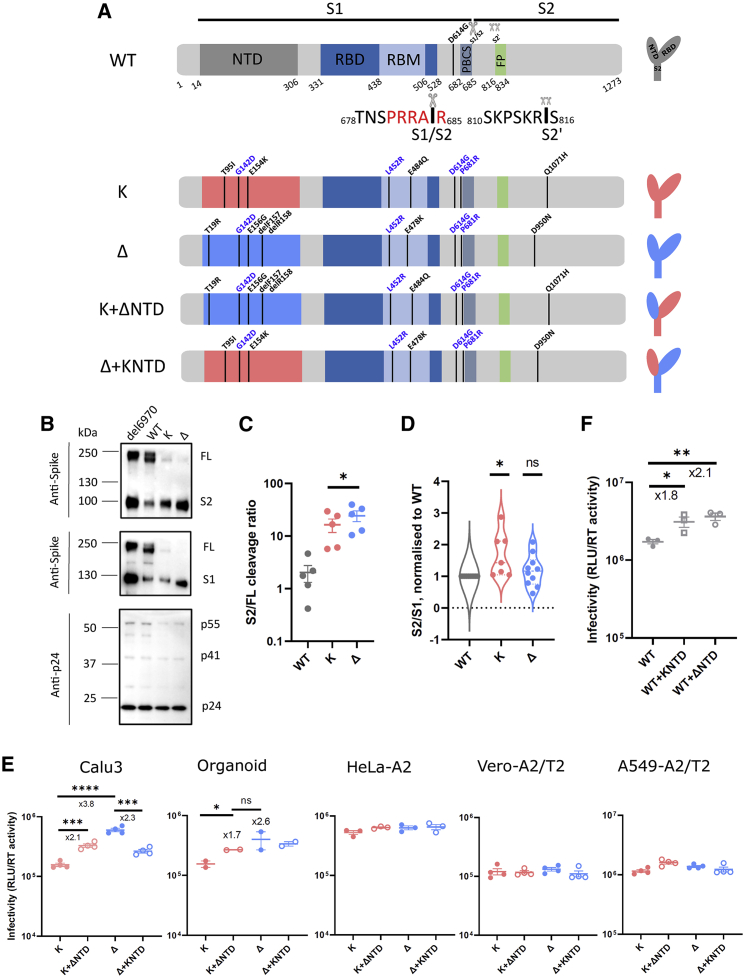
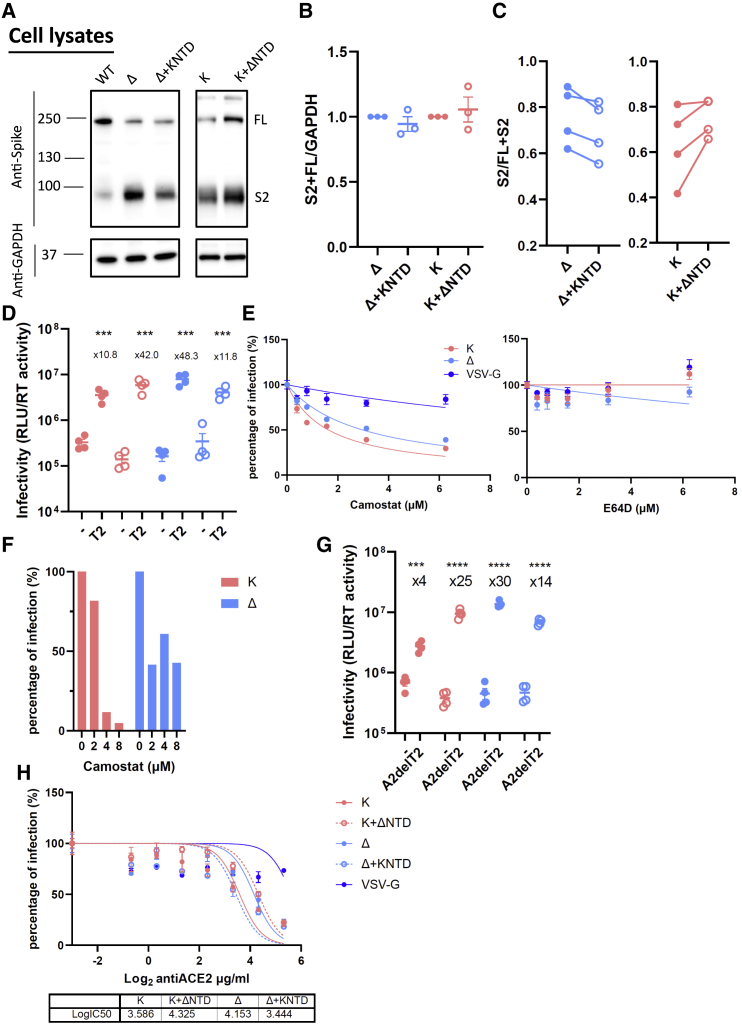
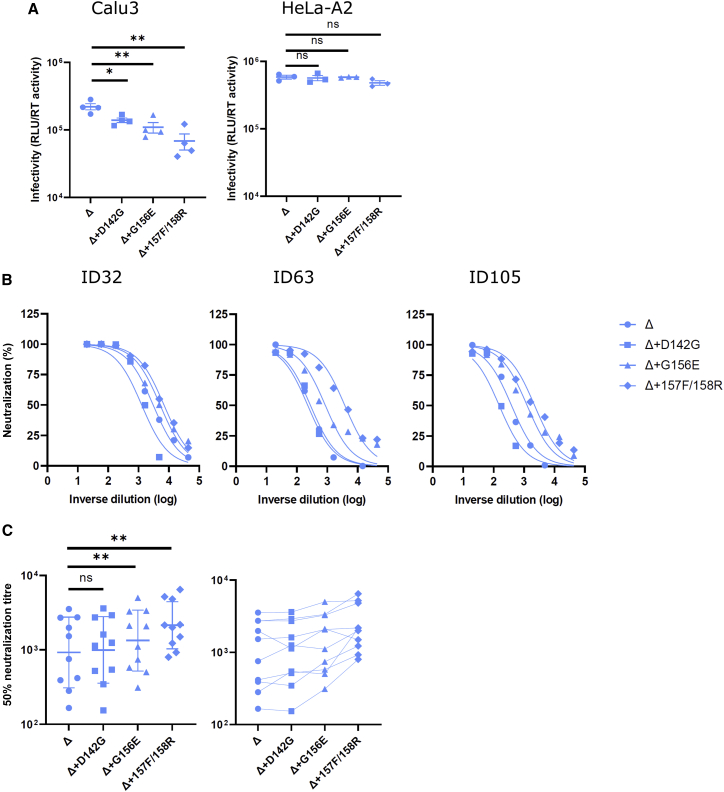
The severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) spike N-terminal domain (NTD) remains poorly characterized despite enrichment of mutations in this region across variants of concern (VOCs). Here, we examine the contribution of the NTD to infection and cell-cell fusion by constructing chimeric spikes bearing B.1.617 lineage (Delta and Kappa variants) NTDs and generating spike pseudotyped lentivirus. We find that the Delta NTD on a Kappa or wild-type (WT) background increases S1/S2 cleavage efficiency and virus entry, specifically in lung cells and airway organoids, through use of TMPRSS2. Delta exhibits increased cell-cell fusogenicity that could be conferred to WT and Kappa spikes by Delta NTD transfer. However, chimeras of Omicron BA.1 and BA.2 spikes with a Delta NTD do not show more efficient TMPRSS2 use or fusogenicity. We conclude that the NTD allosterically modulates S1/S2 cleavage and spike-mediated functions in a spike context-dependent manner, and allosteric interactions may be lost when combining regions from more distantly related VOCs.
Keywords: CP: Microbiology; Delta; NTD; Omicron; Organoid; SARS-CoV-2; TMPRSS2; entry; fusogenicity; spike.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests R.K.G. has received honoraria for consulting and educational activities from Gilead, GSK, Janssen, and Moderna. All the other authors declare no competing interests.
Figures
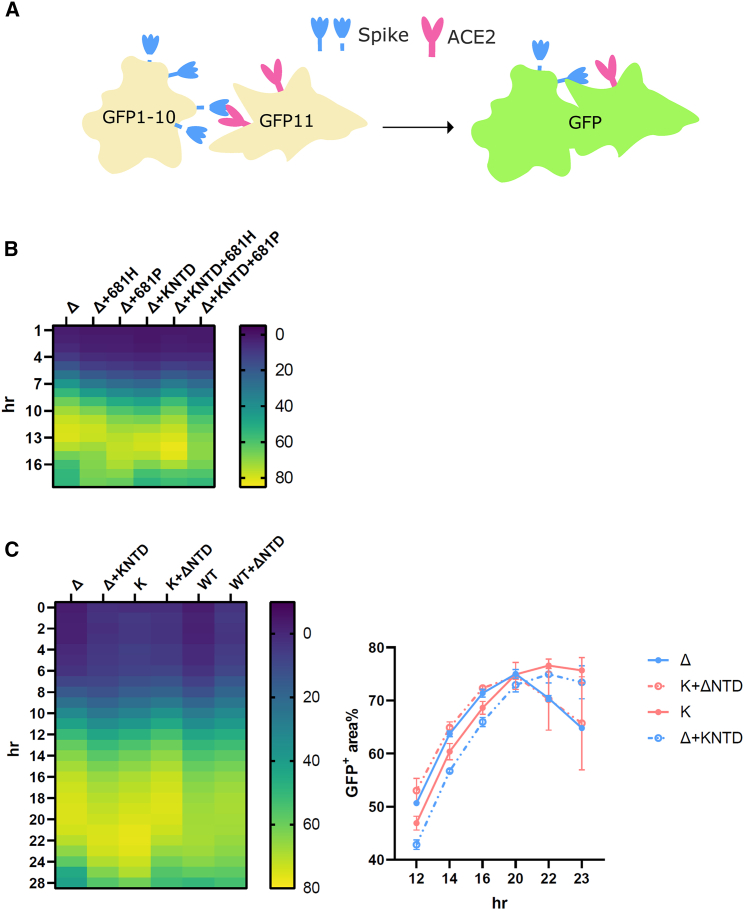
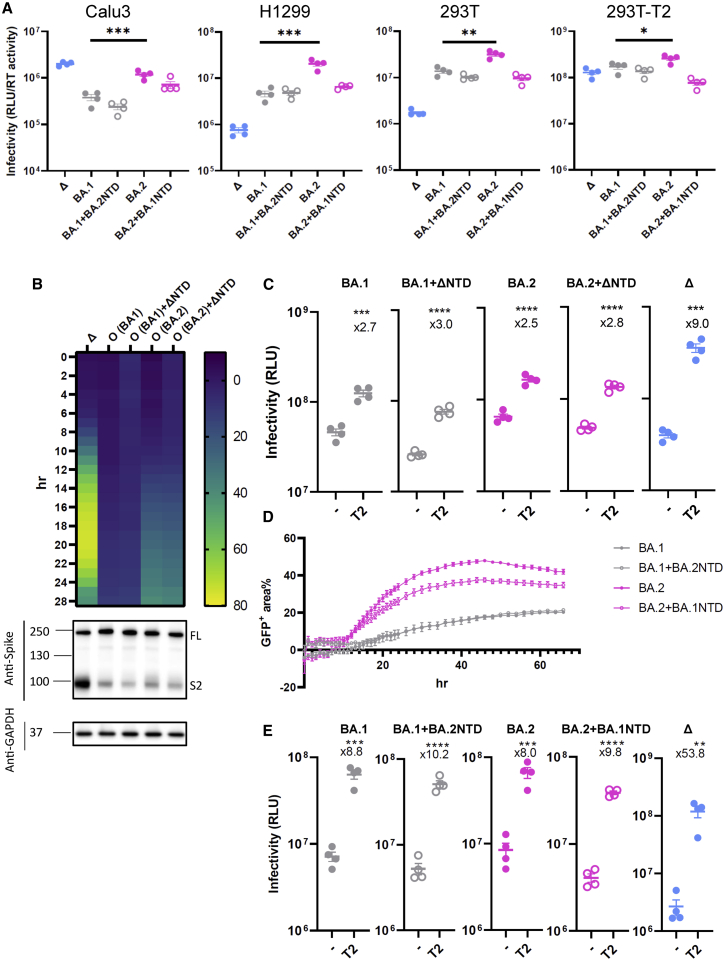
References
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
