

Measurement of ATP utilization in RNA unwinding and RNA chaperone activities by DEAD-box helicase proteins
- PMID: 35965018
- PMCID: PMC10040262
- DOI: 10.1016/bs.mie.2022.04.004
Measurement of ATP utilization in RNA unwinding and RNA chaperone activities by DEAD-box helicase proteins
Abstract
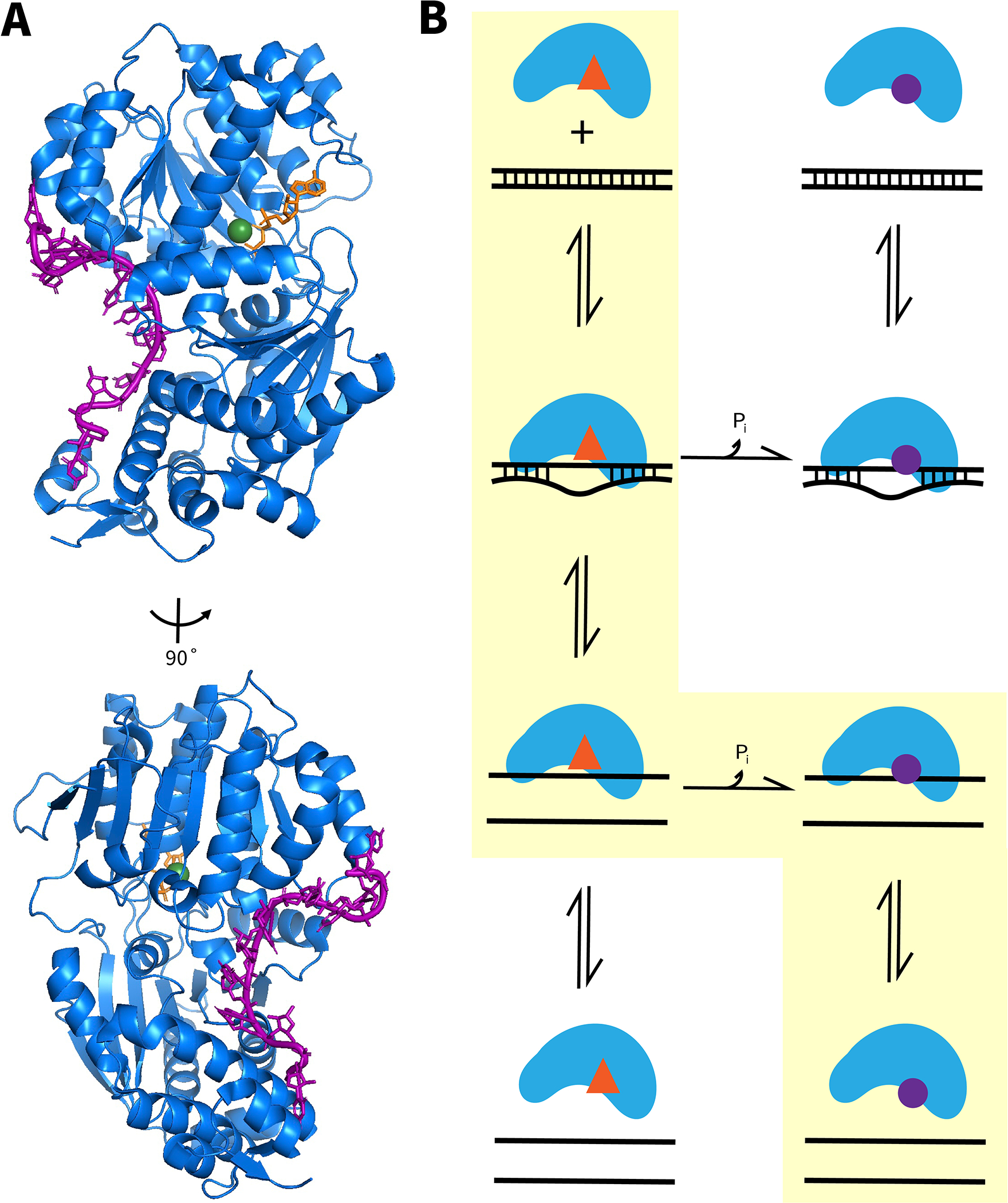
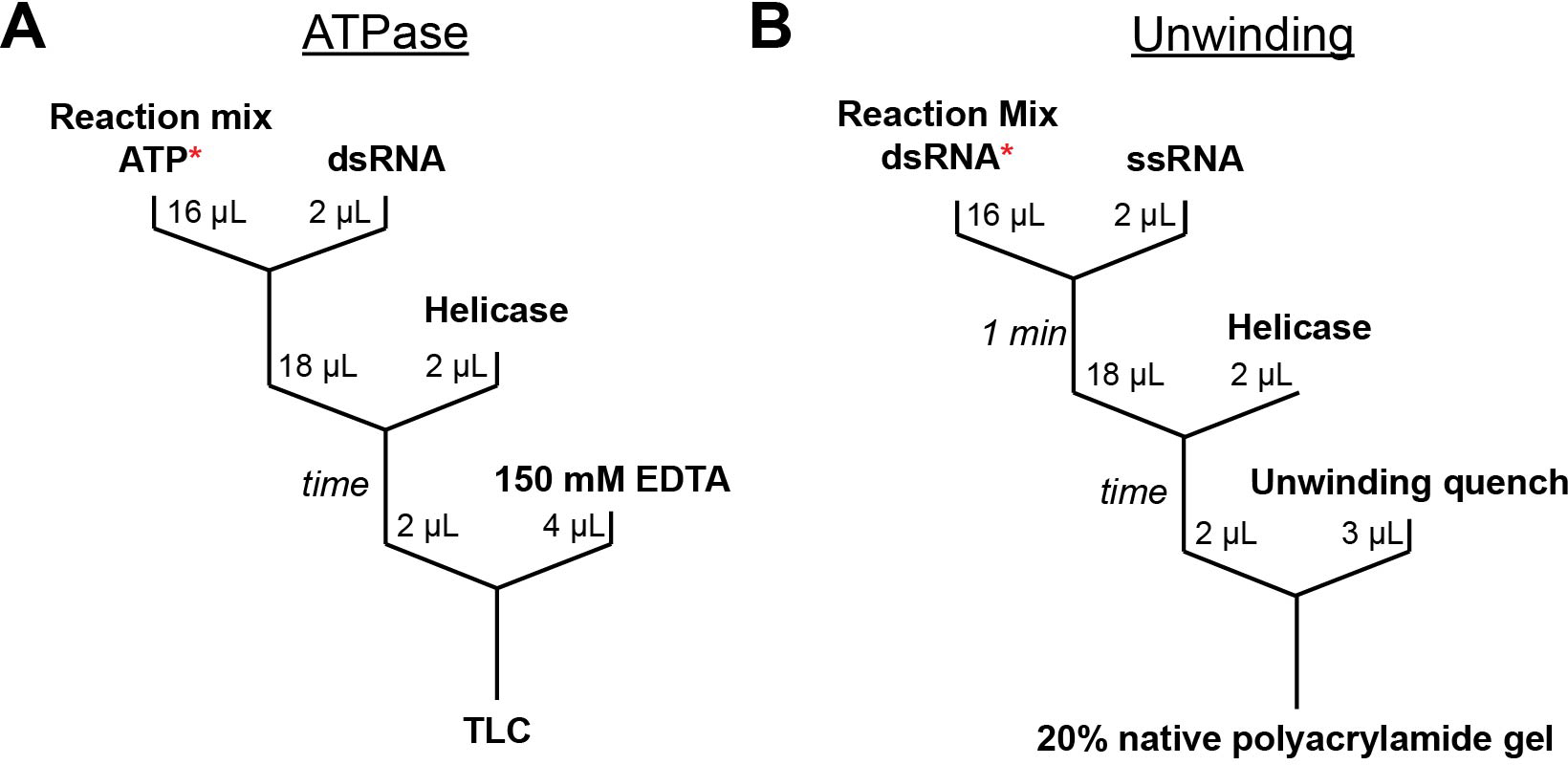
RNA helicase proteins perform coupled reactions in which cycles of ATP binding and hydrolysis are used to drive local unwinding of double-stranded RNA (dsRNA). For some helicases in the ubiquitous DEAD-box family, these local unwinding events are integral to folding transitions in structured RNAs, and thus these helicases function as RNA chaperones. An important measure of the efficiency of the helicase-catalyzed reaction is the ATP utilization value, which represents the average number of ATP molecules hydrolyzed during RNA unwinding or a chaperone-assisted RNA structural rearrangement. Here we outline procedures that can be used to measure the ATP utilization value in RNA unwinding or folding transitions. As an example of an RNA folding transition, we focus on the refolding of the Tetrahymena thermophila group I intron ribozyme from a long-lived misfolded structure to its native structure, and we outline strategies for adapting this assay to other RNA folding transitions. For a simple dsRNA unwinding event, the ATP utilization value provides a measure of the coupling between the ATPase and RNA unwinding activities, and for a complex RNA structural transition it can give insight into the scope of the rearrangement and the efficiency with which the helicase uses the energy from ATPase cycles to promote the rearrangement.
Keywords: ATPase kinetics; CYT-19; DEAD-box protein; Double-stranded RNA; RNA chaperone; RNA helicase; RNA misfolding.
Copyright © 2022 Elsevier Inc. All rights reserved.
Figures



Similar articles
-
ATP utilization by a DEAD-box protein during refolding of a misfolded group I intron ribozyme.J Biol Chem. 2021 Jan-Jun;296:100132. doi: 10.1074/jbc.RA120.015029. Epub 2020 Dec 5. J Biol Chem. 2021. PMID: 33262215 Free PMC article.
-
DEAD-box protein CYT-19 is activated by exposed helices in a group I intron RNA.Proc Natl Acad Sci U S A. 2014 Jul 22;111(29):E2928-36. doi: 10.1073/pnas.1404307111. Epub 2014 Jul 7. Proc Natl Acad Sci U S A. 2014. PMID: 25002474 Free PMC article.
-
Structural basis for RNA-duplex recognition and unwinding by the DEAD-box helicase Mss116p.Nature. 2012 Oct 4;490(7418):121-5. doi: 10.1038/nature11402. Epub 2012 Sep 2. Nature. 2012. PMID: 22940866 Free PMC article.
-
The mechanism of ATP-dependent RNA unwinding by DEAD box proteins.Biol Chem. 2009 Dec;390(12):1237-50. doi: 10.1515/BC.2009.135. Biol Chem. 2009. PMID: 19747077 Review.
-
DEAD-box proteins as RNA helicases and chaperones.Wiley Interdiscip Rev RNA. 2011 Jan-Feb;2(1):135-52. doi: 10.1002/wrna.50. Wiley Interdiscip Rev RNA. 2011. PMID: 21297876 Free PMC article. Review.
Cited by
-
Tracking Native Tetrahymena Ribozyme Folding with Fluorescence.Biochemistry. 2023 Nov 21;62(22):3173-3180. doi: 10.1021/acs.biochem.3c00363. Epub 2023 Nov 1. Biochemistry. 2023. PMID: 37910627 Free PMC article.
References
-
- Atkinson A, & Jack GW (1973). Precipitation of nucleic acids with polyethyleneimine and the chromatography of nucleic acids and proteins on immobilised polyethyleneimine. Biochimica et Biophysica Acta (BBA) - Nucleic Acids and Protein Synthesis, 308(1), 41–52. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
