

Neural Pathway for Gut Feelings: Vagal Interoceptive Feedback From the Gastrointestinal Tract Is a Critical Modulator of Anxiety-like Behavior
- PMID: 35965105
- PMCID: PMC11438499
- DOI: 10.1016/j.biopsych.2022.04.020
Neural Pathway for Gut Feelings: Vagal Interoceptive Feedback From the Gastrointestinal Tract Is a Critical Modulator of Anxiety-like Behavior
Abstract
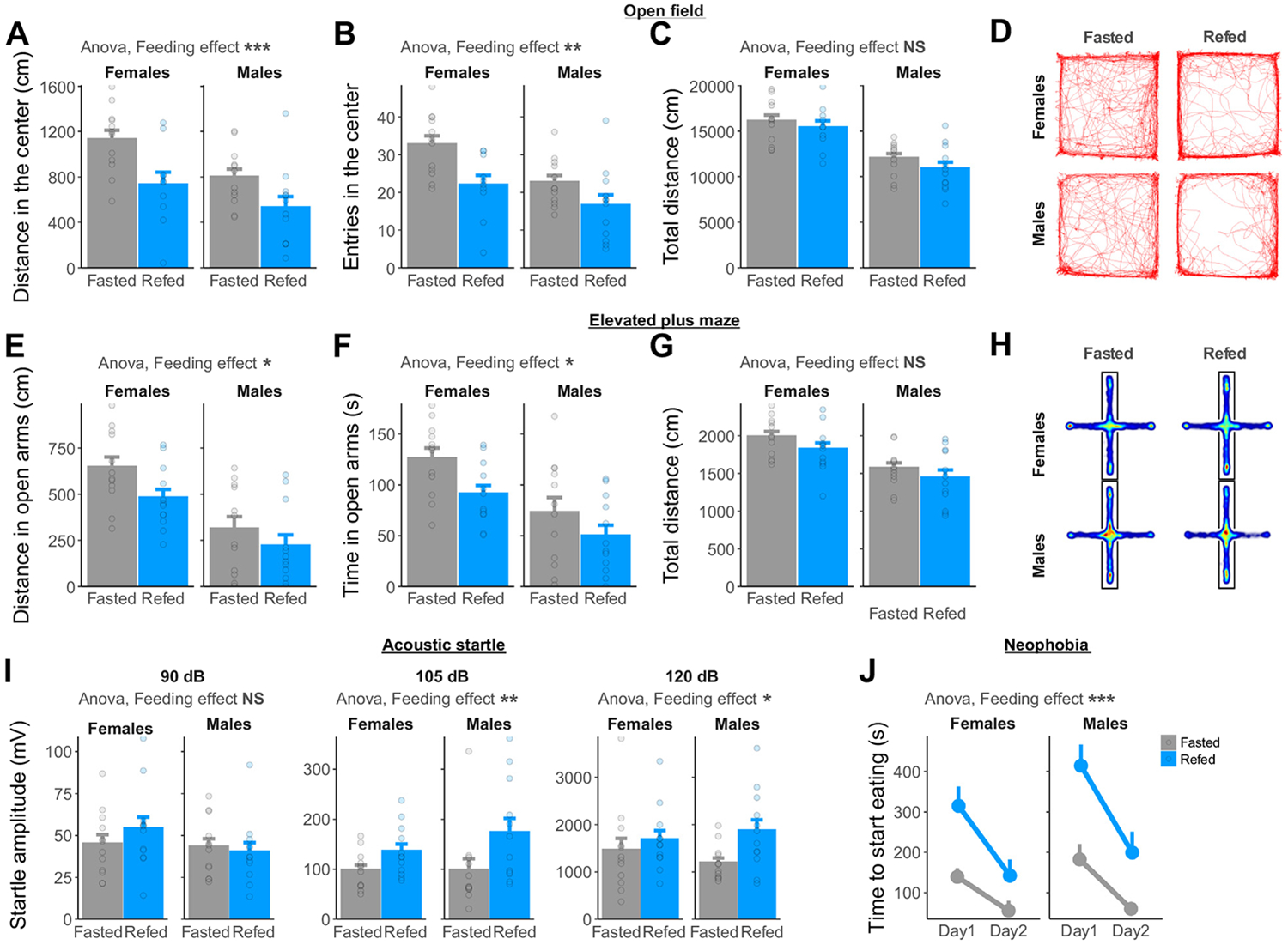
Background: Anxiety disorders are associated with an altered perception of the body's internal state. Therefore, understanding the neuronal basis of interoception can foster novel anxiety therapies. In rodents, the feeding status bidirectionally modulates anxiety-like behavior but how the sensing of gastrointestinal state affects anxiety remains unclear.
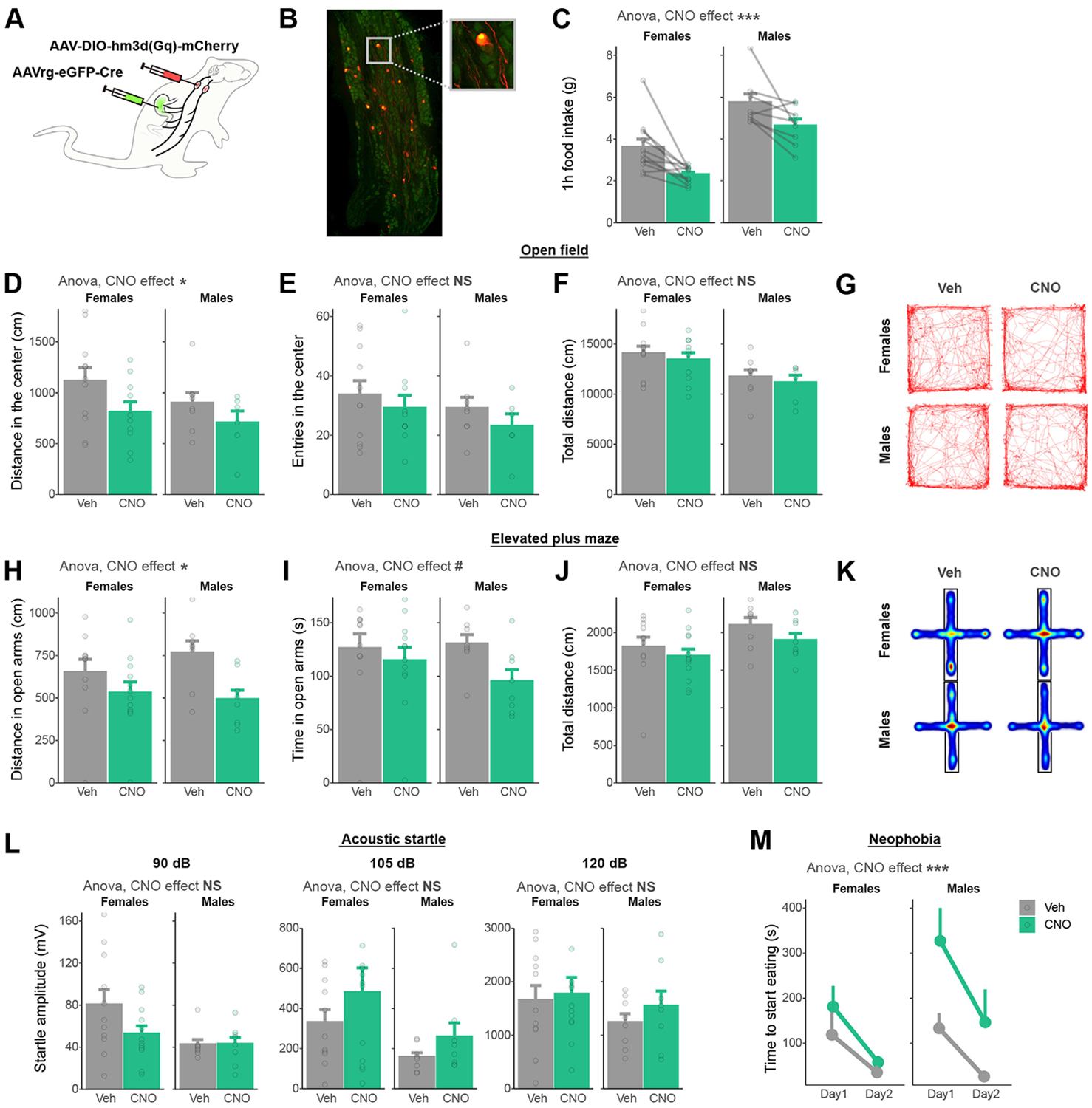
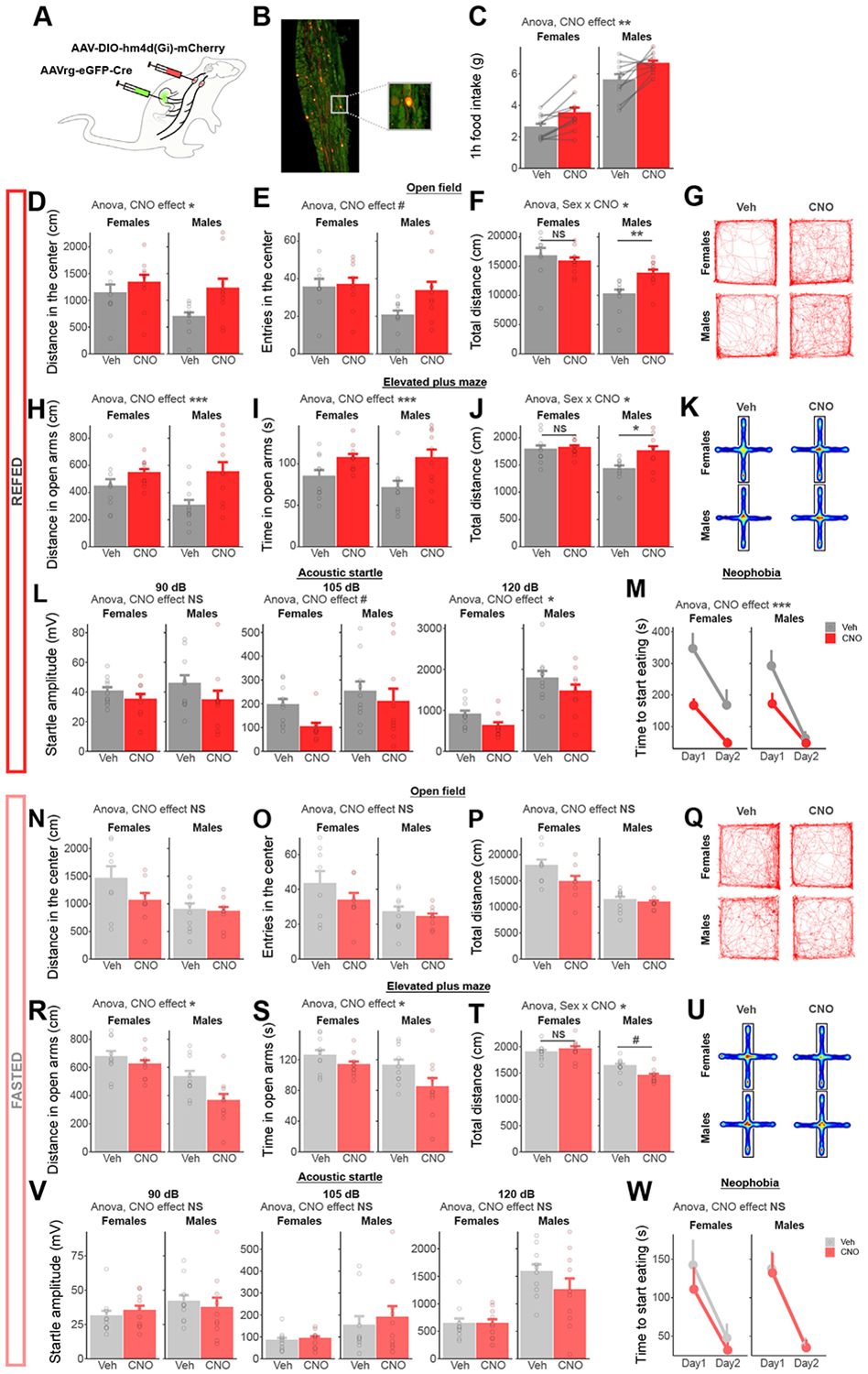
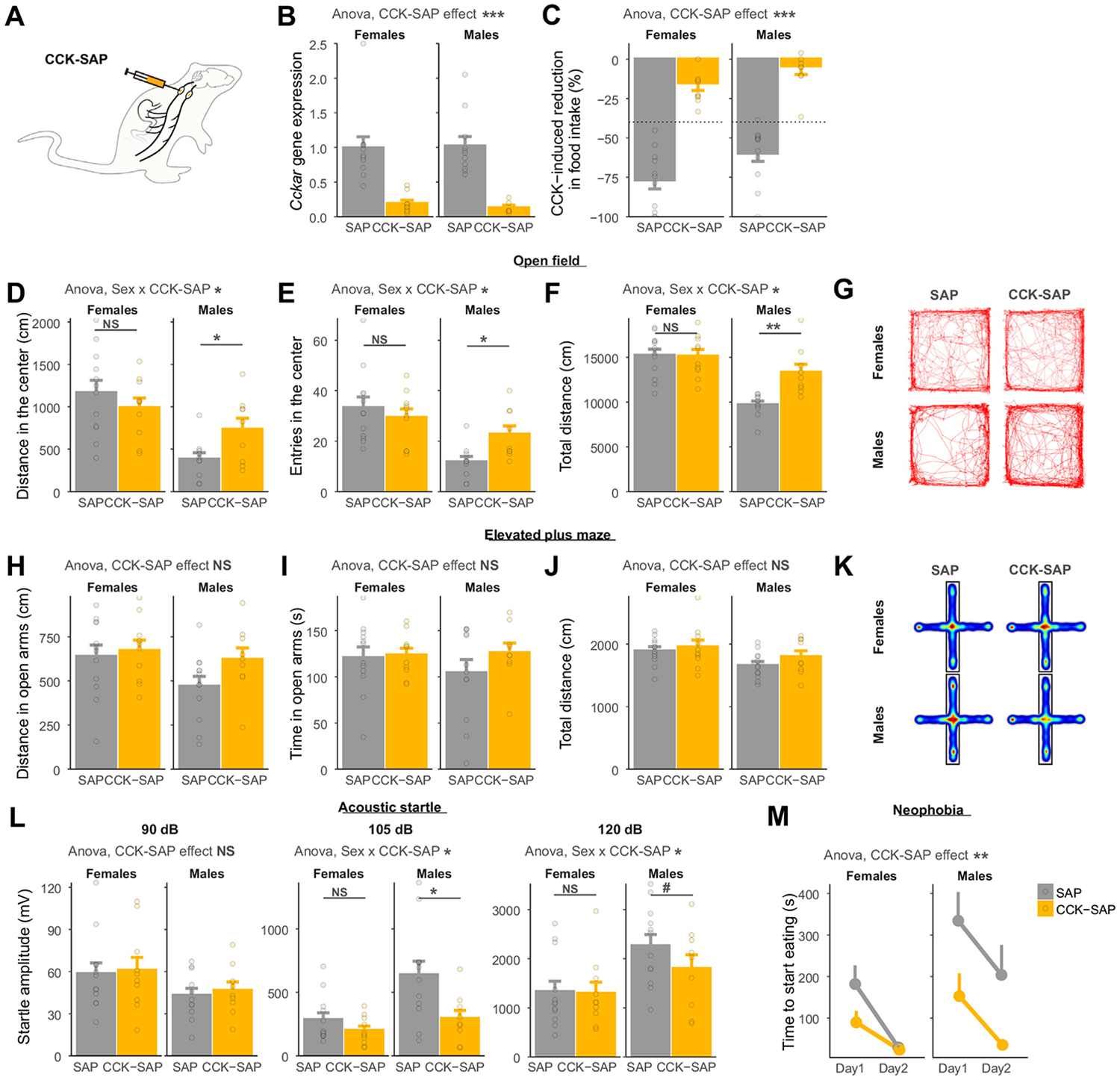
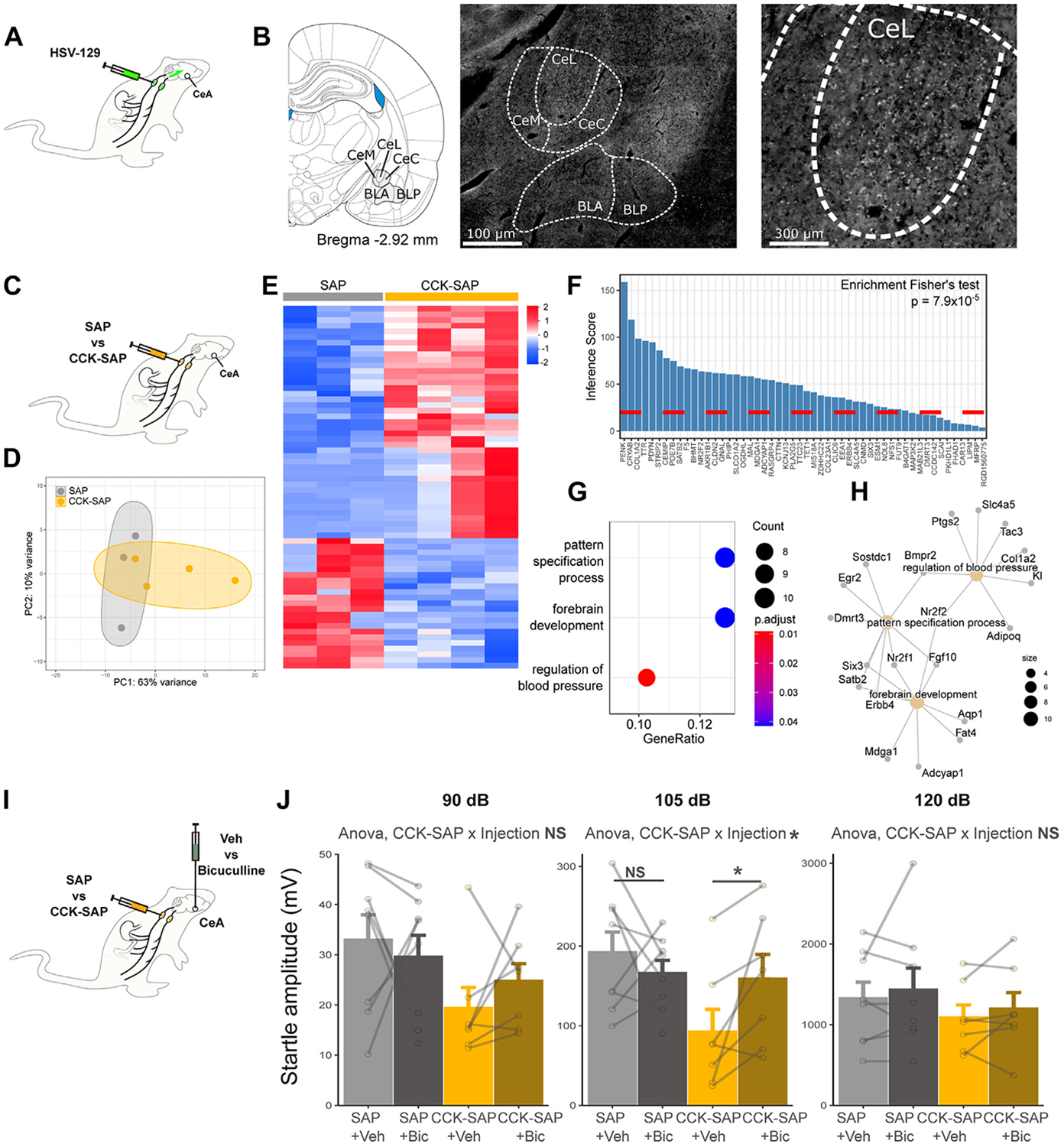
Methods: We combined chemogenetics, neuropharmacology, and behavioral approaches in male and female rats to test whether vagal afferents terminating in the gastrointestinal tract mediate feeding-induced tuning of anxiety. Using saporin-based lesions and transcriptomics, we investigated the chronic impact of this gut-brain circuit on anxiety-like behavior.
Results: Both feeding and selective chemogenetic activation of gut-innervating vagal afferents increased anxiety-like behavior. Conversely, chemogenetic inhibition blocked the increase in anxiety-like behavior induced by feeding. Using a selective saporin-based lesion, we demonstrate that the loss of gut-innervating vagal afferent signaling chronically reduces anxiety-like behavior in male rats but not in female rats. We next identify a vagal circuit that connects the gut to the central nucleus of the amygdala, using anterograde transsynaptic tracing from the nodose ganglia. Lesion of this gut-brain vagal circuit modulated the central amygdala transcriptome in both sexes but selectively affected a network of GABA (gamma-aminobutyric acid)-related genes only in males, suggesting a potentiation of inhibitory control. Blocking GABAergic signaling in the central amygdala re-established normal anxiety levels in male rats.
Conclusions: Vagal sensory signals from the gastrointestinal tract are critical for baseline and feeding-induced tuning of anxiety via the central amygdala in rats. Our results suggest vagal gut-brain signaling as a target to normalize interoception in anxiety disorders.
Keywords: Amygdala; Anxiety; Gut-brains axis; Interoception; Sex differences; Vagus nerve.
Copyright © 2022 Society of Biological Psychiatry. Published by Elsevier Inc. All rights reserved.
Figures
Comment in
-
Behavioral Consequences of a Rumbling Tummy: Fasting Alters Emotional State via the Vagus Nerve.Biol Psychiatry. 2022 Nov 1;92(9):690-692. doi: 10.1016/j.biopsych.2022.08.006. Biol Psychiatry. 2022. PMID: 36202543 No abstract available.
References
-
- GBD 2016 Disease and Injury Incidence and Prevalence Collaborators (2017): Global, Regional, and National Incidence, Prevalence, and Years lived with disability for 328 diseases and injuries for 195 countries, 1990–2016: A systematic analysis for the Global Burden of Disease Study 2016. Lancet 390:1211–1259. - PMC - PubMed
-
- Domschke K, Stevens S, Pfleiderer B, Gerlach AL (2010): Interoceptive sensitivity in anxiety and anxiety disorders: An overview and integration of neurobiological findings. Clin Psychol Rev 30:1–11. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
