

Exercise training enhances muscle mitochondrial metabolism in diet-resistant obesity
- PMID: 35965199
- PMCID: PMC9482931
- DOI: 10.1016/j.ebiom.2022.104192
Exercise training enhances muscle mitochondrial metabolism in diet-resistant obesity
Abstract
Background: Current paradigms for predicting weight loss in response to energy restriction have general validity but a subset of individuals fail to respond adequately despite documented diet adherence. Patients in the bottom 20% for rate of weight loss following a hypocaloric diet (diet-resistant) have been found to have less type I muscle fibres and lower skeletal muscle mitochondrial function, leading to the hypothesis that physical exercise may be an effective treatment when diet alone is inadequate. In this study, we aimed to assess the efficacy of exercise training on mitochondrial function in women with obesity with a documented history of minimal diet-induced weight loss.
Methods: From over 5000 patient records, 228 files were reviewed to identify baseline characteristics of weight loss response from women with obesity who were previously classified in the top or bottom 20% quintiles based on rate of weight loss in the first 6 weeks during which a 900 kcal/day meal replacement was consumed. A subset of 20 women with obesity were identified based on diet-resistance (n=10) and diet sensitivity (n=10) to undergo a 6-week supervised, progressive, combined aerobic and resistance exercise intervention.
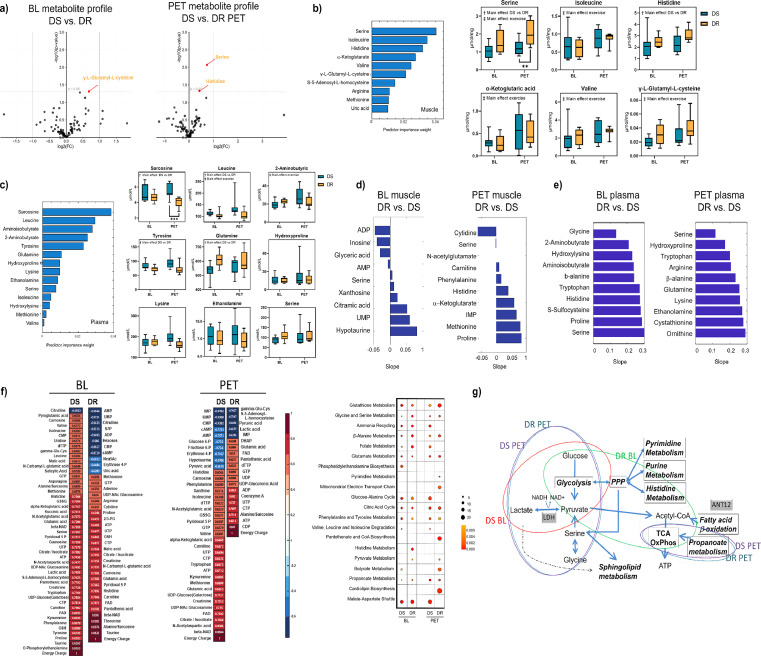
Findings: Diet-sensitive women had lower baseline adiposity, higher fasting insulin and triglycerides, and a greater number of ATP-III criteria for metabolic syndrome. Conversely in diet-resistant women, the exercise intervention improved body composition, skeletal muscle mitochondrial content and metabolism, with minimal effects in diet-sensitive women. In-depth analyses of muscle metabolomes revealed distinct group- and intervention- differences, including lower serine-associated sphingolipid synthesis in diet-resistant women following exercise training.
Interpretation: Exercise preferentially enhances skeletal muscle metabolism and improves body composition in women with a history of minimal diet-induced weight loss. These clinical and metabolic mechanism insights move the field towards better personalised approaches for the treatment of distinct obesity phenotypes.
Funding: Canadian Institutes of Health Research (CIHR-INMD and FDN-143278; CAN-163902; CIHR PJT-148634).
Keywords: Exercise; Metabolomics; Mitochondria; Mitochondrial supercomplexes; Muscle physiology; Obesity; Serine; Sphingolipids; Uncoupling; Weight loss.
Copyright © 2022 The Author(s). Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no conflicts of interest.
Figures




Similar articles
-
Mitochondrial capacity in skeletal muscle is not stimulated by weight loss despite increases in insulin action and decreases in intramyocellular lipid content.Diabetes. 2008 Apr;57(4):987-94. doi: 10.2337/db07-1429. Epub 2008 Feb 5. Diabetes. 2008. PMID: 18252894 Clinical Trial.
-
Skeletal muscle metabolism in overweight and post-overweight women: an isometric exercise study using (31)P magnetic resonance spectroscopy.Int J Obes Relat Metab Disord. 2001 Sep;25(9):1309-15. doi: 10.1038/sj.ijo.0801673. Int J Obes Relat Metab Disord. 2001. PMID: 11571592
-
Decreased mitochondrial proton leak and reduced expression of uncoupling protein 3 in skeletal muscle of obese diet-resistant women.Diabetes. 2002 Aug;51(8):2459-66. doi: 10.2337/diabetes.51.8.2459. Diabetes. 2002. PMID: 12145158
-
Sphingolipids as a Culprit of Mitochondrial Dysfunction in Insulin Resistance and Type 2 Diabetes.Front Endocrinol (Lausanne). 2021 Mar 18;12:635175. doi: 10.3389/fendo.2021.635175. eCollection 2021. Front Endocrinol (Lausanne). 2021. PMID: 33815291 Free PMC article. Review.
-
The role of weight loss and exercise in correcting skeletal muscle mitochondrial abnormalities in obesity, diabetes and aging.Mol Cell Endocrinol. 2013 Oct 15;379(1-2):30-4. doi: 10.1016/j.mce.2013.06.018. Epub 2013 Jun 20. Mol Cell Endocrinol. 2013. PMID: 23792186 Review.
Cited by
-
Muscle fiber phenotype: a culprit of abnormal metabolism and function in skeletal muscle of humans with obesity.Am J Physiol Endocrinol Metab. 2023 Dec 1;325(6):E723-E733. doi: 10.1152/ajpendo.00190.2023. Epub 2023 Oct 25. Am J Physiol Endocrinol Metab. 2023. PMID: 37877797 Free PMC article. Review.
-
Impact of combined exercise on blood DNA methylation and physical health in older women with obesity.PLoS One. 2024 Dec 16;19(12):e0315250. doi: 10.1371/journal.pone.0315250. eCollection 2024. PLoS One. 2024. PMID: 39680552 Free PMC article.
-
The Impact of Diet and Physical Activity on Fat-to-Lean Mass Ratio.Nutrients. 2023 Dec 20;16(1):19. doi: 10.3390/nu16010019. Nutrients. 2023. PMID: 38201847 Free PMC article.
-
The relationship between muscle mass and fat content in body composition and non-alcoholic fatty liver disease in the Chinese general population: a cross-sectional study.Front Med (Lausanne). 2024 Jun 10;11:1384366. doi: 10.3389/fmed.2024.1384366. eCollection 2024. Front Med (Lausanne). 2024. PMID: 38915765 Free PMC article.
-
Mitochondrial transplantation: adaptive bio-enhancement.Cell Death Dis. 2025 Jul 1;16(1):473. doi: 10.1038/s41419-025-07643-8. Cell Death Dis. 2025. PMID: 40595522 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
