

Bps polysaccharide of Bordetella pertussis resists antimicrobial peptides by functioning as a dual surface shield and decoy and converts Escherichia coli into a respiratory pathogen
- PMID: 35969621
- PMCID: PMC9410548
- DOI: 10.1371/journal.ppat.1010764
Bps polysaccharide of Bordetella pertussis resists antimicrobial peptides by functioning as a dual surface shield and decoy and converts Escherichia coli into a respiratory pathogen
Abstract
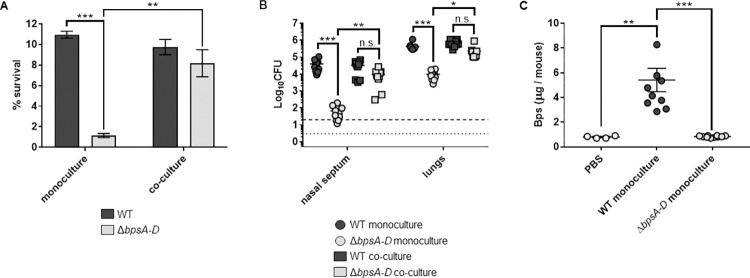
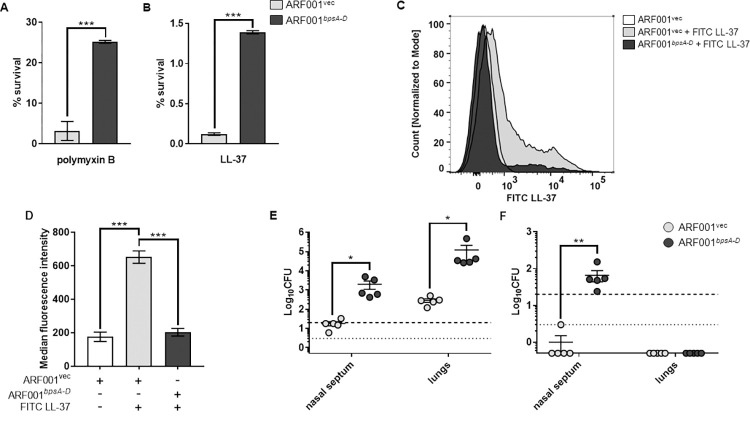
Infections and disease caused by the obligate human pathogen Bordetella pertussis (Bp) are increasing, despite widespread vaccinations. The current acellular pertussis vaccines remain ineffective against nasopharyngeal colonization, carriage, and transmission. In this work, we tested the hypothesis that Bordetella polysaccharide (Bps), a member of the poly-β-1,6-N-acetyl-D-glucosamine (PNAG/PGA) family of polysaccharides promotes respiratory tract colonization of Bp by resisting killing by antimicrobial peptides (AMPs). Genetic deletion of the bpsA-D locus, as well as treatment with the specific glycoside hydrolase Dispersin B, increased susceptibility to AMP-mediated killing. Bps was found to be both cell surface-associated and released during laboratory growth and mouse infections. Addition of bacterial supernatants containing Bps and purified Bps increased B. pertussis resistance to AMPs. By utilizing ELISA, immunoblot and flow cytometry assays, we show that Bps functions as a dual surface shield and decoy. Co-inoculation of C57BL/6J mice with a Bps-proficient strain enhanced respiratory tract survival of the Bps-deficient strain. In combination, the presented results highlight the critical role of Bps as a central driver of B. pertussis pathogenesis. Heterologous production of Bps in a non-pathogenic E. coli K12 strain increased AMP resistance in vitro, and augmented bacterial survival and pathology in the mouse respiratory tract. These studies can serve as a foundation for other PNAG/PGA polysaccharides and for the development of an effective Bp vaccine that includes Bps.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
The Bordetella pertussis Bps polysaccharide enhances lung colonization by conferring protection from complement-mediated killing.Cell Microbiol. 2014 Jul;16(7):1105-18. doi: 10.1111/cmi.12264. Epub 2014 Feb 13. Cell Microbiol. 2014. PMID: 24438122 Free PMC article.
-
The Bps polysaccharide of Bordetella pertussis promotes colonization and biofilm formation in the nose by functioning as an adhesin.Mol Microbiol. 2010 Sep;77(6):1439-55. doi: 10.1111/j.1365-2958.2010.07297.x. Epub 2010 Jul 30. Mol Microbiol. 2010. PMID: 20633227 Free PMC article.
-
The protein BpsB is a poly-β-1,6-N-acetyl-D-glucosamine deacetylase required for biofilm formation in Bordetella bronchiseptica.J Biol Chem. 2015 Sep 11;290(37):22827-40. doi: 10.1074/jbc.M115.672469. Epub 2015 Jul 22. J Biol Chem. 2015. PMID: 26203190 Free PMC article.
-
Bordetella pertussis and pertactin-deficient clinical isolates: lessons for pertussis vaccines.Expert Rev Vaccines. 2014 Sep;13(9):1135-46. doi: 10.1586/14760584.2014.932254. Epub 2014 Jun 23. Expert Rev Vaccines. 2014. PMID: 24953157 Review.
-
Molecular Epidemiology of Bordetella pertussis.Adv Exp Med Biol. 2019;1183:19-33. doi: 10.1007/5584_2019_402. Adv Exp Med Biol. 2019. PMID: 31342459 Review.
Cited by
-
A Comprehensive Review of Recent Research into the Effects of Antimicrobial Peptides on Biofilms-January 2020 to September 2023.Antibiotics (Basel). 2024 Apr 9;13(4):343. doi: 10.3390/antibiotics13040343. Antibiotics (Basel). 2024. PMID: 38667019 Free PMC article. Review.
-
Development of carbohydrate based next-generation anti-pertussis vaccines.Bioorg Med Chem. 2022 Nov 15;74:117066. doi: 10.1016/j.bmc.2022.117066. Epub 2022 Oct 17. Bioorg Med Chem. 2022. PMID: 36283250 Free PMC article. Review.
-
Extracellular defense of bacteria against antimicrobial peptides.J Bacteriol. 2025 Aug 21;207(8):e0016625. doi: 10.1128/jb.00166-25. Epub 2025 Aug 1. J Bacteriol. 2025. PMID: 40748075 Free PMC article. Review.
-
Protective Activity and Safety of Experimental Acellular Pertussis Vaccines Based on Antigenic Complexes Isolated from Biofilm and Planktonic Cultures of Bordetella pertussis.Bull Exp Biol Med. 2024 Jul;177(3):349-352. doi: 10.1007/s10517-024-06187-9. Epub 2024 Aug 10. Bull Exp Biol Med. 2024. PMID: 39126548
-
Highlights of the 14th International Bordetella Symposium.mSphere. 2025 Jun 25;10(6):e0018925. doi: 10.1128/msphere.00189-25. Epub 2025 May 16. mSphere. 2025. PMID: 40377335 Free PMC article. Review.
References
-
- Bowden KE, Weigand MR, Peng Y, Cassiday PK, Sammons S, Knipe K, et al.. Genome Structural Diversity among 31 Bordetella pertussis Isolates from Two Recent U.S. Whooping Cough Statewide Epidemics. mSphere. 2016;1(3). Epub 2016/06/16. doi: 10.1128/mSphere.00036-16 ; PubMed Central PMCID: PMC4888882. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
