

Two distinct ways to form long-term object recognition memory during sleep and wakefulness
- PMID: 35969775
- PMCID: PMC9407643
- DOI: 10.1073/pnas.2203165119
Two distinct ways to form long-term object recognition memory during sleep and wakefulness
Abstract
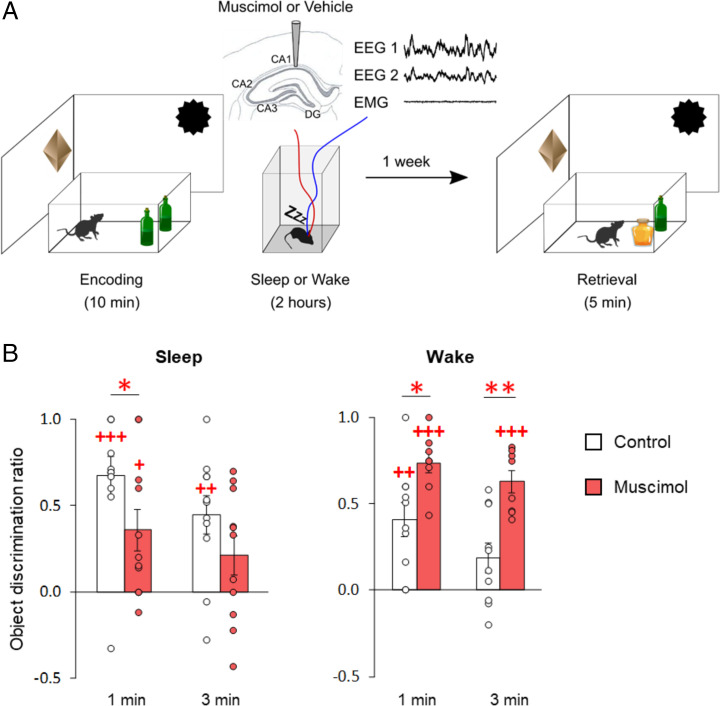
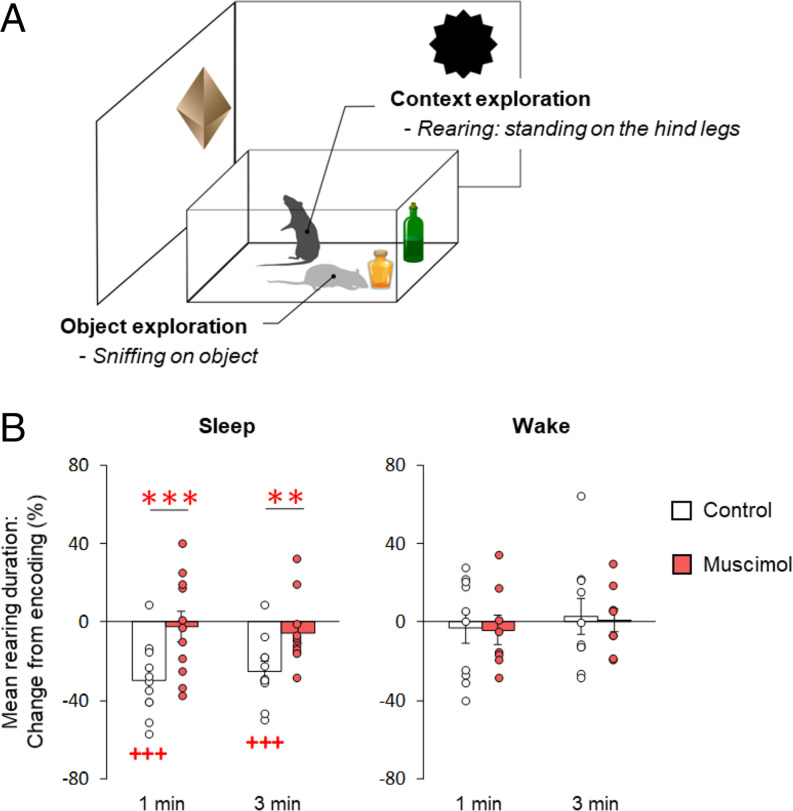
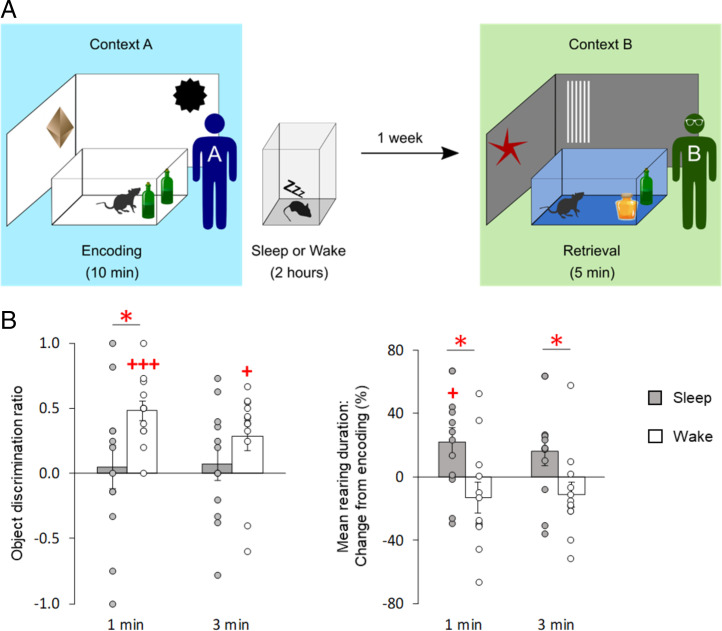
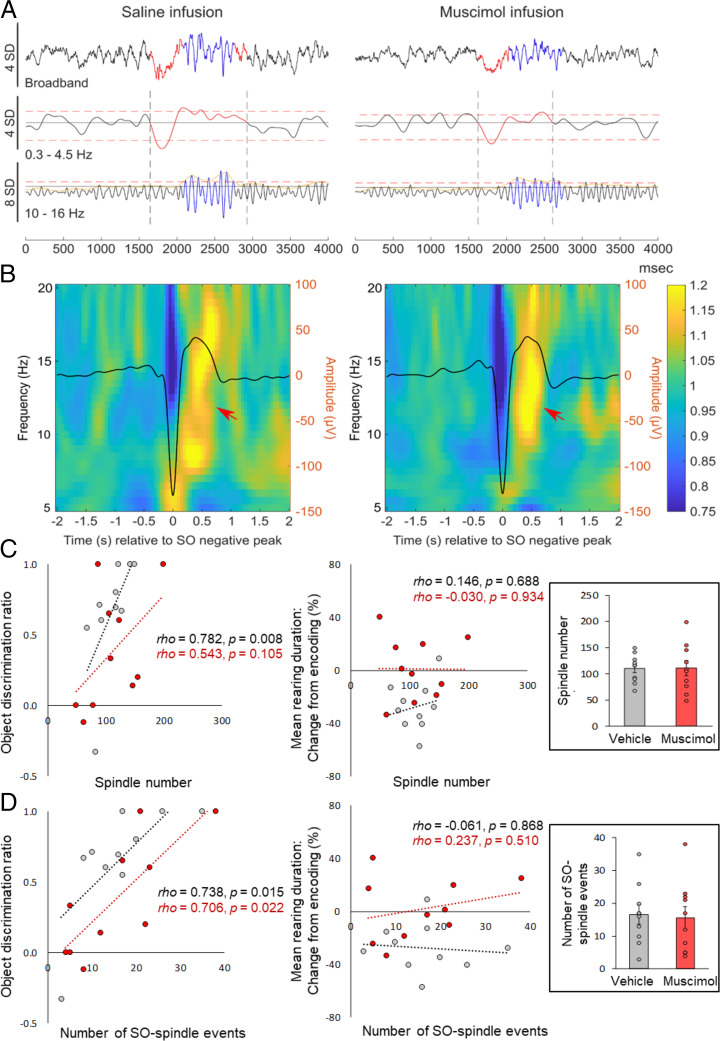
Memory consolidation is promoted by sleep. However, there is also evidence for consolidation into long-term memory during wakefulness via processes that preferentially affect nonhippocampal representations. We compared, in rats, the effects of 2-h postencoding periods of sleep and wakefulness on the formation of long-term memory for objects and their associated environmental contexts. We employed a novel-object recognition (NOR) task, using object exploration and exploratory rearing as behavioral indicators of these memories. Remote recall testing (after 1 wk) confirmed significant long-term NOR memory under both conditions, with NOR memory after sleep predicted by the occurrence of EEG spindle-slow oscillation coupling. Rats in the sleep group decreased their exploratory rearing at recall testing, revealing successful recall of the environmental context. By contrast, rats that stayed awake after encoding showed equally high levels of rearing upon remote testing as during encoding, indicating that context memory was lost. Disruption of hippocampal function during the postencoding interval (by muscimol administration) suppressed long-term NOR memory together with context memory formation when animals slept, but enhanced NOR memory when they were awake during this interval. Testing remote recall in a context different from that during encoding impaired NOR memory in the sleep condition, while exploratory rearing was increased. By contrast, NOR memory in the wake rats was preserved and actually superior to that after sleep. Our findings indicate two distinct modes of long-term memory formation: Sleep consolidation is hippocampus dependent and implicates event-context binding, whereas wake consolidation is impaired by hippocampal activation and strengthens context-independent representations.
Keywords: memory consolidation; novel-object recognition memory; sleep; wake.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
The hippocampus is crucial for forming non-hippocampal long-term memory during sleep.Nature. 2018 Dec;564(7734):109-113. doi: 10.1038/s41586-018-0716-8. Epub 2018 Nov 14. Nature. 2018. PMID: 30429612
-
Sleep in Humans Stabilizes Pattern Separation Performance.J Neurosci. 2017 Dec 13;37(50):12238-12246. doi: 10.1523/JNEUROSCI.1189-17.2017. Epub 2017 Nov 8. J Neurosci. 2017. PMID: 29118106 Free PMC article.
-
Hippocampal corticosterone impairs memory consolidation during sleep but improves consolidation in the wake state.Hippocampus. 2014 May;24(5):510-5. doi: 10.1002/hipo.22266. Epub 2014 Mar 17. Hippocampus. 2014. PMID: 24596244 Free PMC article.
-
Sleep-A brain-state serving systems memory consolidation.Neuron. 2023 Apr 5;111(7):1050-1075. doi: 10.1016/j.neuron.2023.03.005. Neuron. 2023. PMID: 37023710 Review.
-
Mechanisms of systems memory consolidation during sleep.Nat Neurosci. 2019 Oct;22(10):1598-1610. doi: 10.1038/s41593-019-0467-3. Epub 2019 Aug 26. Nat Neurosci. 2019. PMID: 31451802 Review.
Cited by
-
How sleep shapes what we remember-and forget.Proc Natl Acad Sci U S A. 2023 Jan 10;120(2):e2220275120. doi: 10.1073/pnas.2220275120. Epub 2023 Jan 3. Proc Natl Acad Sci U S A. 2023. PMID: 36595692 Free PMC article. No abstract available.
-
Rearing Behavior as Indicator of Spatial Novelty and Memory in Developing Rats.Eur J Neurosci. 2025 Jun;61(12):e70162. doi: 10.1111/ejn.70162. Eur J Neurosci. 2025. PMID: 40538152 Free PMC article.
-
Effect of Aging and a Dual Orexin Receptor Antagonist on Sleep Architecture and Non-REM Oscillations Including an REM Behavior Disorder Phenotype in the PS19 Mouse Model of Tauopathy.J Neurosci. 2023 Jun 21;43(25):4738-4749. doi: 10.1523/JNEUROSCI.1828-22.2023. Epub 2023 May 25. J Neurosci. 2023. PMID: 37230765 Free PMC article.
-
Post-conditioning sleep deprivation facilitates delay and trace fear memory extinction.Mol Brain. 2024 Nov 29;17(1):90. doi: 10.1186/s13041-024-01163-w. Mol Brain. 2024. PMID: 39614358 Free PMC article.
-
Multi-region processing during sleep for memory and cognition.Proc Jpn Acad Ser B Phys Biol Sci. 2025;101(3):107-128. doi: 10.2183/pjab.101.008. Proc Jpn Acad Ser B Phys Biol Sci. 2025. PMID: 40074337 Free PMC article. Review.
References
-
- Stickgold R., Sleep-dependent memory consolidation. Nature 437, 1272–1278 (2005). - PubMed
-
- Inostroza M., Binder S., Born J., Sleep-dependency of episodic-like memory consolidation in rats. Behav. Brain Res. 237, 15–22 (2013). - PubMed
-
- Ishikawa H., Yamada K., Pavlides C., Ichitani Y., Sleep deprivation impairs spontaneous object-place but not novel-object recognition in rats. Neurosci. Lett. 580, 114–118 (2014). - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
