

Allosteric inhibitors targeting the calmodulin-PIP2 interface of SK4 K+ channels for atrial fibrillation treatment
- PMID: 35969786
- PMCID: PMC9407317
- DOI: 10.1073/pnas.2202926119
Allosteric inhibitors targeting the calmodulin-PIP2 interface of SK4 K+ channels for atrial fibrillation treatment
Abstract
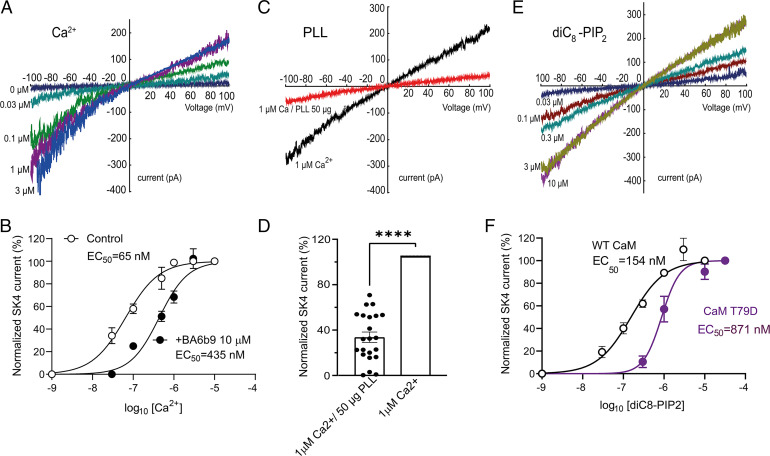
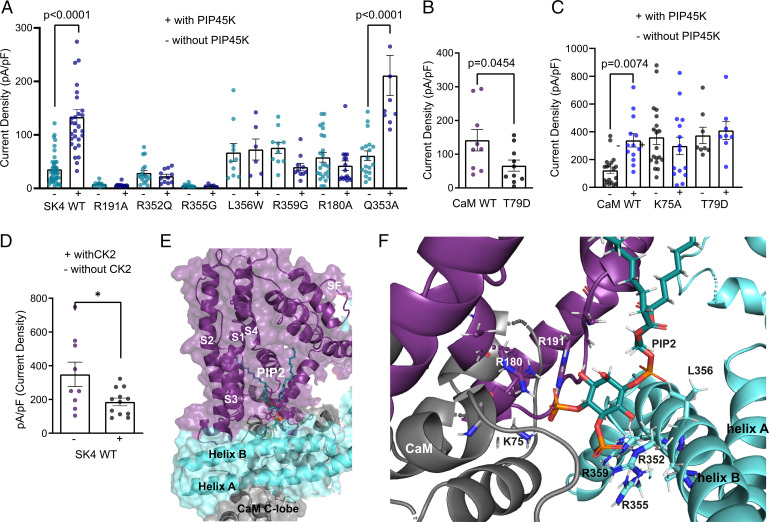
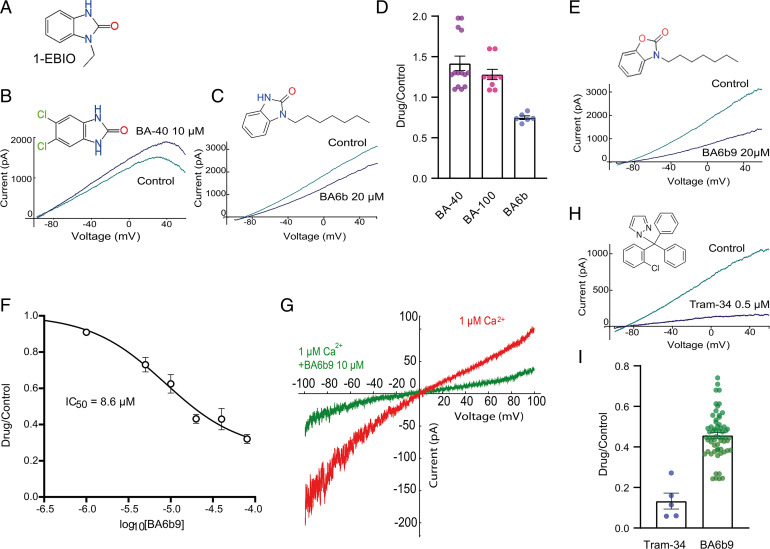
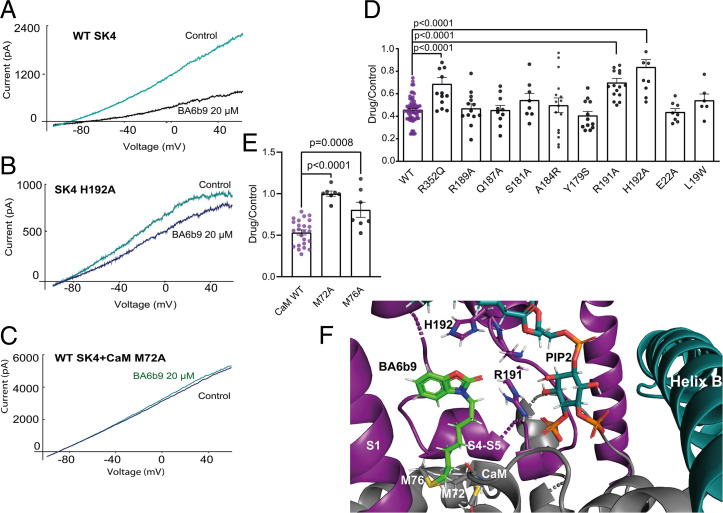
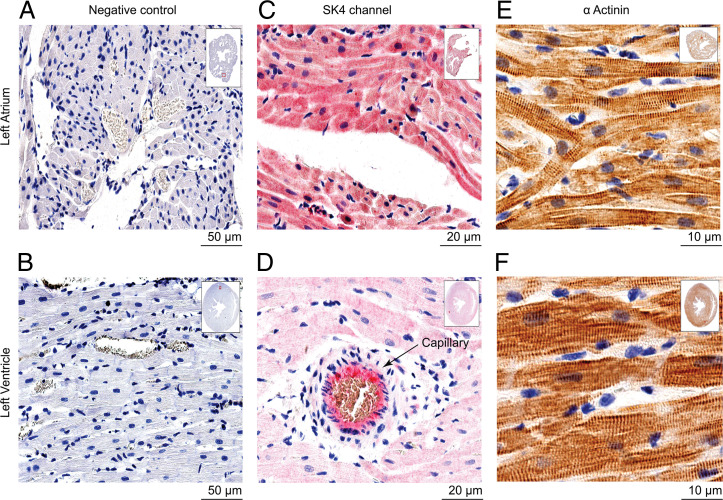
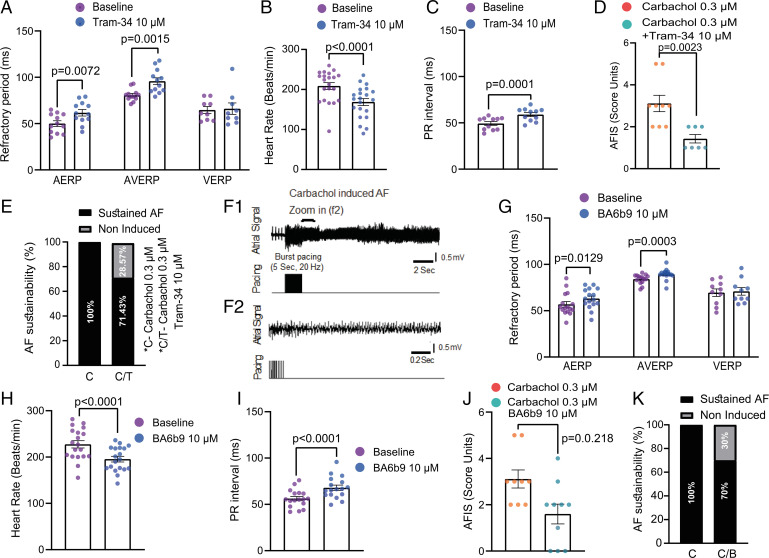
The Ca2+-activated SK4 K+ channel is gated by Ca2+-calmodulin (CaM) and is expressed in immune cells, brain, and heart. A cryoelectron microscopy (cryo-EM) structure of the human SK4 K+ channel recently revealed four CaM molecules per channel tetramer, where the apo CaM C-lobe and the holo CaM N-lobe interact with the proximal carboxyl terminus and the linker S4-S5, respectively, to gate the channel. Here, we show that phosphatidylinositol 4-5 bisphosphate (PIP2) potently activates SK4 channels by docking to the boundary of the CaM-binding domain. An allosteric blocker, BA6b9, was designed to act to the CaM-PIP2-binding domain, a previously untargeted region of SK4 channels, at the interface of the proximal carboxyl terminus and the linker S4-S5. Site-directed mutagenesis, molecular docking, and patch-clamp electrophysiology indicate that BA6b9 inhibits SK4 channels by interacting with two specific residues, Arg191 and His192 in the linker S4-S5, not conserved in SK1-SK3 subunits, thereby conferring selectivity and preventing the Ca2+-CaM N-lobe from properly interacting with the channel linker region. Immunohistochemistry of the SK4 channel protein in rat hearts showed a widespread expression in the sarcolemma of atrial myocytes, with a sarcomeric striated Z-band pattern, and a weaker occurrence in the ventricle but a marked incidence at the intercalated discs. BA6b9 significantly prolonged atrial and atrioventricular effective refractory periods in rat isolated hearts and reduced atrial fibrillation induction ex vivo. Our work suggests that inhibition of SK4 K+ channels by targeting drugs to the CaM-PIP2-binding domain provides a promising anti-arrhythmic therapy.
Keywords: KCa3.1; PIP2; atrial fibrillation; calmodulin; potassium channel.
Conflict of interest statement
Competing interest statement: The authors declare a competing interest. There is a pending patent application filed for the new chemical entities disclosed in the manuscript.
Figures
Similar articles
-
Competition of calcified calmodulin N lobe and PIP2 to an LQT mutation site in Kv7.1 channel.Proc Natl Acad Sci U S A. 2017 Jan 31;114(5):E869-E878. doi: 10.1073/pnas.1612622114. Epub 2017 Jan 17. Proc Natl Acad Sci U S A. 2017. PMID: 28096388 Free PMC article.
-
The SK4 channel allosteric blocker, BA6b9, reduces atrial fibrillation substrate in rats with reduced ejection fraction.PNAS Nexus. 2024 May 12;3(5):pgae192. doi: 10.1093/pnasnexus/pgae192. eCollection 2024 May. PNAS Nexus. 2024. PMID: 38783894 Free PMC article.
-
Activation mechanism of a human SK-calmodulin channel complex elucidated by cryo-EM structures.Science. 2018 May 4;360(6388):508-513. doi: 10.1126/science.aas9466. Science. 2018. PMID: 29724949 Free PMC article.
-
Channelopathy of small- and intermediate-conductance Ca2+-activated K+ channels.Acta Pharmacol Sin. 2023 Feb;44(2):259-267. doi: 10.1038/s41401-022-00935-1. Epub 2022 Jun 17. Acta Pharmacol Sin. 2023. PMID: 35715699 Free PMC article. Review.
-
Atomistic Insights of Calmodulin Gating of Complete Ion Channels.Int J Mol Sci. 2020 Feb 14;21(4):1285. doi: 10.3390/ijms21041285. Int J Mol Sci. 2020. PMID: 32075037 Free PMC article. Review.
Cited by
-
Atrial fibroblast-derived exosomal miR-21 upregulate myocardial KCa3.1 via the PI3K-Akt pathway during rapid pacing.Heliyon. 2024 Jun 14;10(13):e33059. doi: 10.1016/j.heliyon.2024.e33059. eCollection 2024 Jul 15. Heliyon. 2024. PMID: 39040331 Free PMC article.
-
Intracellular acidity impedes KCa3.1 activation by Riluzole and SKA-31.Front Pharmacol. 2024 Apr 4;15:1380655. doi: 10.3389/fphar.2024.1380655. eCollection 2024. Front Pharmacol. 2024. PMID: 38638868 Free PMC article.
-
Atomistic mechanisms of the regulation of small-conductance Ca2+-activated K+ channel (SK2) by PIP2.Proc Natl Acad Sci U S A. 2024 Sep 24;121(39):e2318900121. doi: 10.1073/pnas.2318900121. Epub 2024 Sep 17. Proc Natl Acad Sci U S A. 2024. PMID: 39288178 Free PMC article.
-
Characterization and Chemical Synthesis of Cm39 (α-KTx 4.8): A Scorpion Toxin That Inhibits Voltage-Gated K+ Channel KV1.2 and Small- and Intermediate-Conductance Ca2+-Activated K+ Channels KCa2.2 and KCa3.1.Toxins (Basel). 2023 Jan 5;15(1):41. doi: 10.3390/toxins15010041. Toxins (Basel). 2023. PMID: 36668861 Free PMC article.
-
IK Channel-Independent Effects of Clotrimazole and Senicapoc on Cancer Cells Viability and Migration.Int J Mol Sci. 2023 Nov 14;24(22):16285. doi: 10.3390/ijms242216285. Int J Mol Sci. 2023. PMID: 38003471 Free PMC article.
References
-
- Berkefeld H., Fakler B., Schulte U., Ca2+-activated K+ channels: From protein complexes to function. Physiol. Rev. 90, 1437–1459 (2010). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
