

Prolonged FOS activity disrupts a global myogenic transcriptional program by altering 3D chromatin architecture in primary muscle progenitor cells
- PMID: 35971133
- PMCID: PMC9377060
- DOI: 10.1186/s13395-022-00303-x
Prolonged FOS activity disrupts a global myogenic transcriptional program by altering 3D chromatin architecture in primary muscle progenitor cells
Abstract
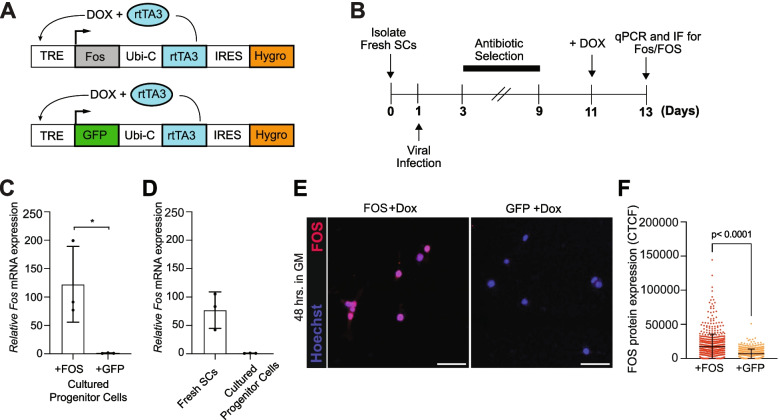
Background: The AP-1 transcription factor, FBJ osteosarcoma oncogene (FOS), is induced in adult muscle satellite cells (SCs) within hours following muscle damage and is required for effective stem cell activation and muscle repair. However, why FOS is rapidly downregulated before SCs enter cell cycle as progenitor cells (i.e., transiently expressed) remains unclear. Further, whether boosting FOS levels in the proliferating progeny of SCs can enhance their myogenic properties needs further evaluation.
Methods: We established an inducible, FOS expression system to evaluate the impact of persistent FOS activity in muscle progenitor cells ex vivo. We performed various assays to measure cellular proliferation and differentiation, as well as uncover changes in RNA levels and three-dimensional (3D) chromatin interactions.
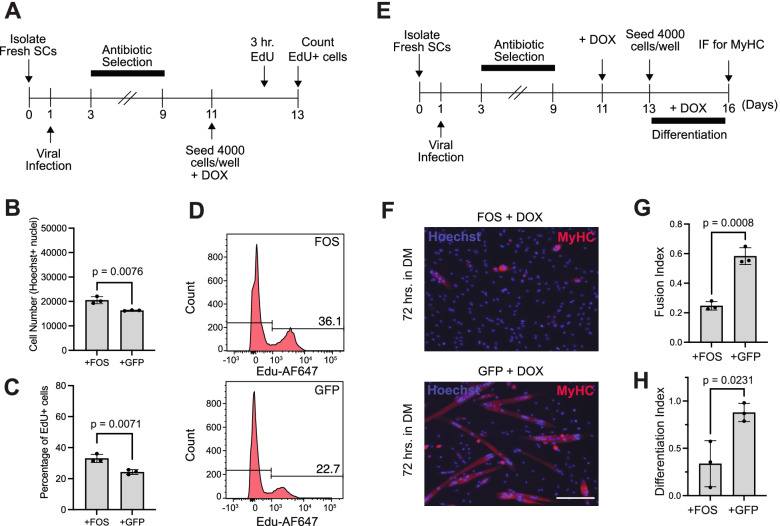
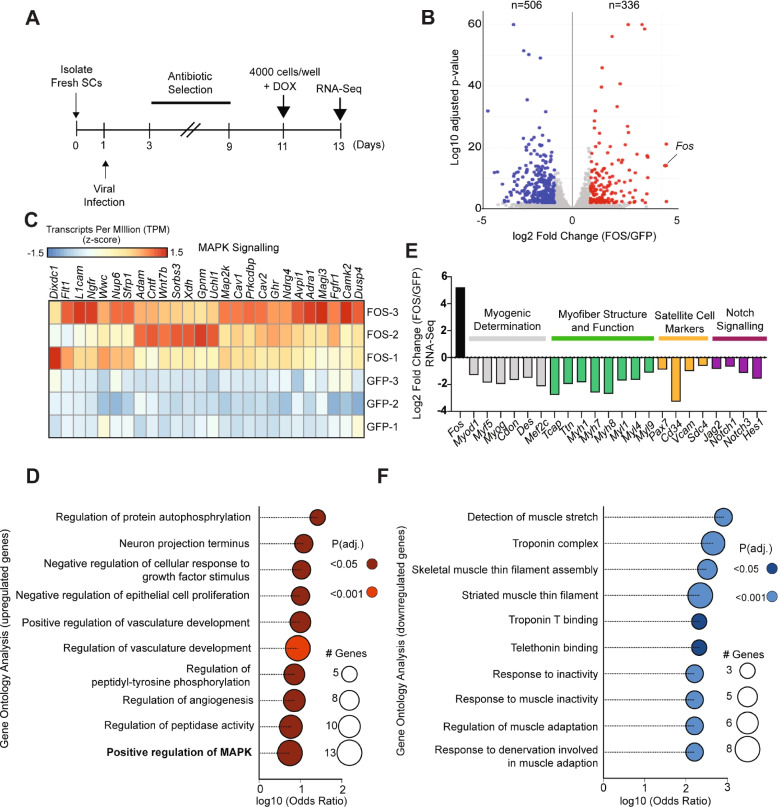
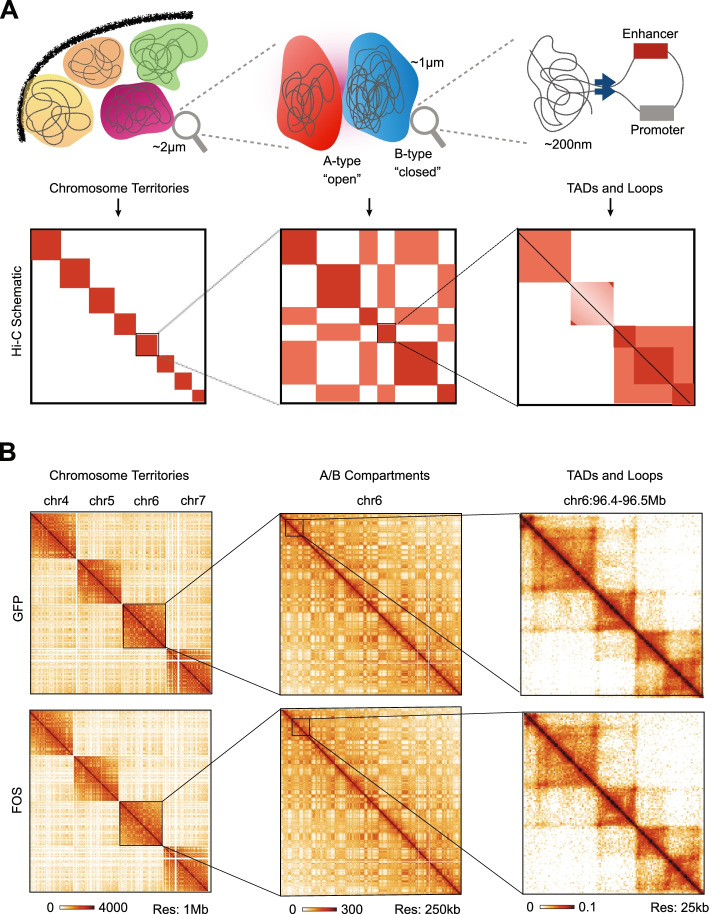
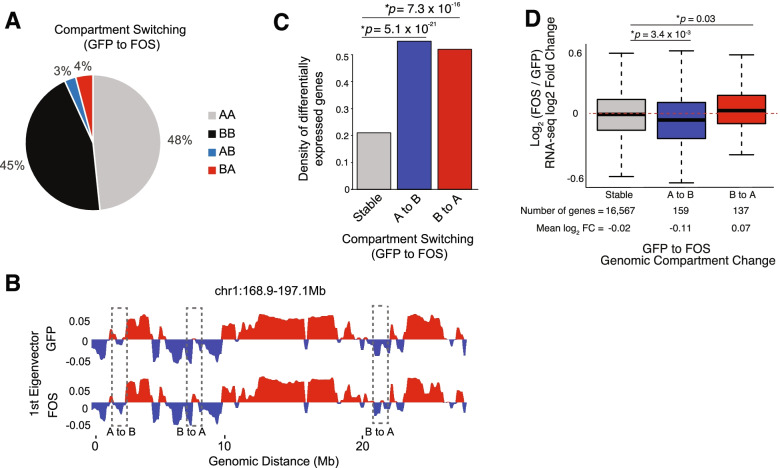
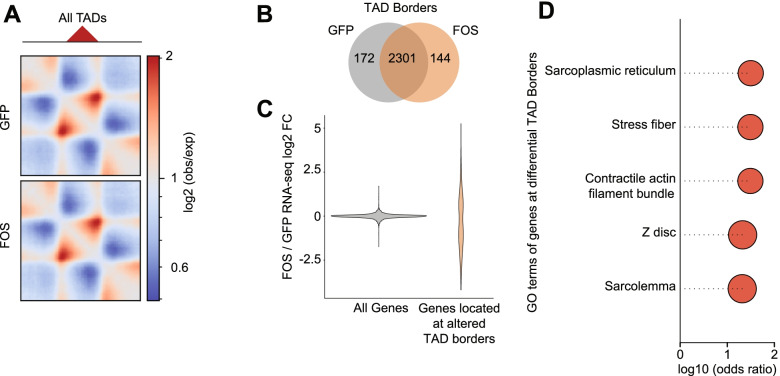
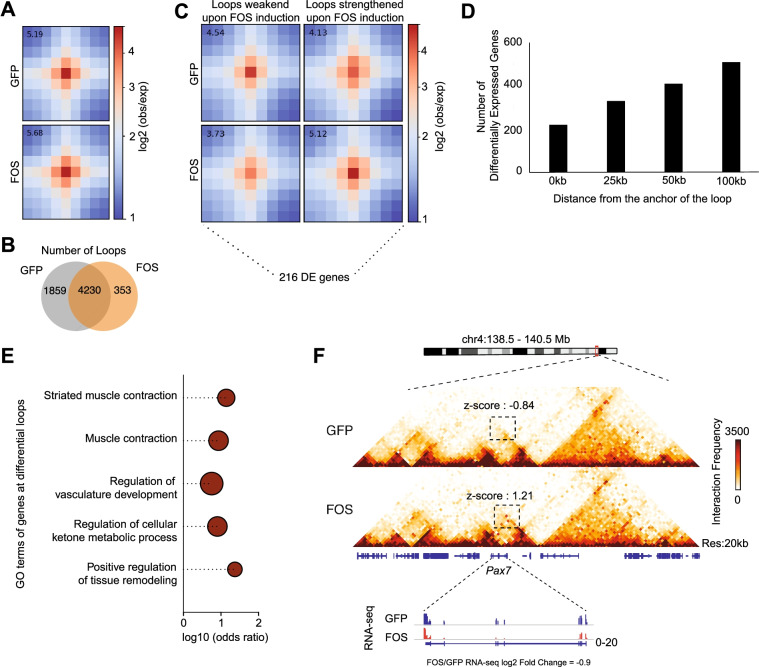
Results: Persistent FOS activity in primary muscle progenitor cells severely antagonizes their ability to differentiate and form myotubes within the first 2 weeks in culture. RNA-seq analysis revealed that ectopic FOS activity in muscle progenitor cells suppressed a global pro-myogenic transcriptional program, while activating a stress-induced, mitogen-activated protein kinase (MAPK) transcriptional signature. Additionally, we observed various FOS-dependent, chromosomal re-organization events in A/B compartments, topologically associated domains (TADs), and genomic loops near FOS-regulated genes.
Conclusions: Our results suggest that elevated FOS activity in recently activated muscle progenitor cells perturbs cellular differentiation by altering the 3D chromosome organization near critical pro-myogenic genes. This work highlights the crucial importance of tightly controlling FOS expression in the muscle lineage and suggests that in states of chronic stress or disease, persistent FOS activity in muscle precursor cells may disrupt the muscle-forming process.
Keywords: AP-1; FOS; Hi-C; Muscle progenitor cells; Muscle satellite cells; Myogenic differentiation; Topologically associated domains (TADs), gene loops.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
