

A SlCLV3-SlWUS module regulates auxin and ethylene homeostasis in low light-induced tomato flower abscission
- PMID: 35972422
- PMCID: PMC9614458
- DOI: 10.1093/plcell/koac254
A SlCLV3-SlWUS module regulates auxin and ethylene homeostasis in low light-induced tomato flower abscission
Abstract
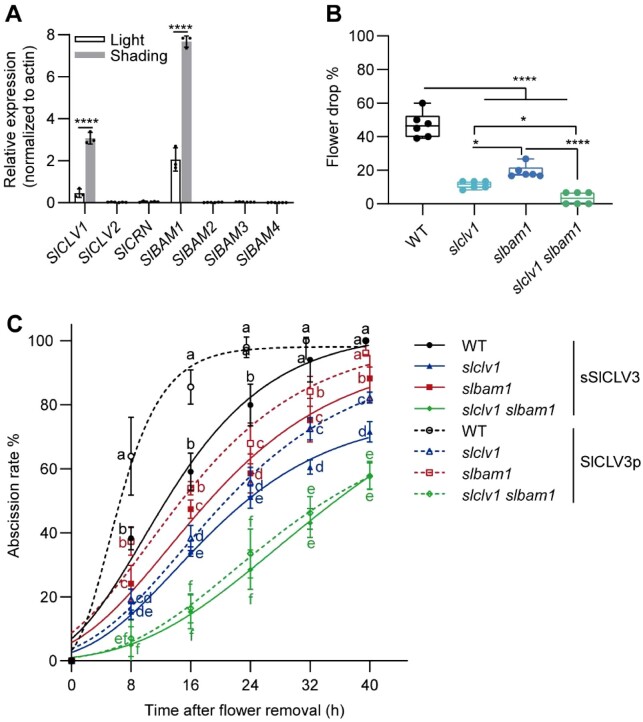
Premature abscission of flowers and fruits triggered by low light stress can severely reduce crop yields. However, the underlying molecular mechanism of this organ abscission is not fully understood. Here, we show that a gene (SlCLV3) encoding CLAVATA3 (CLV3), a peptide hormone that regulates stem cell fate in meristems, is highly expressed in the pedicel abscission zone (AZ) in response to low light in tomato (Solanum lycopersicum). SlCLV3 knockdown and knockout lines exhibit delayed low light-induced flower drop. The receptor kinases SlCLV1 and BARELY ANY MERISTEM1 function in the SlCLV3 peptide-induced low light response in the AZ to decrease expression of the transcription factor gene WUSCHEL (SlWUS). DNA affinity purification sequencing identified the transcription factor genes KNOX-LIKE HOMEDOMAIN PROTEIN1 (SlKD1) and FRUITFULL2 (SlFUL2) as SlWUS target genes. Our data reveal that low light reduces SlWUS expression, resulting in higher SlKD1 and SlFUL2 expression in the AZ, thereby perturbing the auxin response gradient and causing increased ethylene production, eventually leading to the initiation of abscission. These results demonstrate that the SlCLV3-SlWUS signaling pathway plays a central role in low light-induced abscission by affecting auxin and ethylene homeostasis.
© American Society of Plant Biologists 2022. All rights reserved. For permissions, please email: journals.permissions@oup.com.
Figures







References
-
- Brand U, Fletcher JC, Hobe M, Meyerowitz EM, Simon R (2000) Dependence of stem cell fate in Arabidopsis on a feedback loop regulated by CLV3 activity. Science 289: 617–619 - PubMed
-
- Busch W, Miotk A, Ariel FD, Zhao Z, Forner J, Daum G, Suzaki T, Schuster C, Schultheiss SJ, Leibfried A (2010) Transcriptional control of a plant stem cell niche. Dev Cell 18: 841–853 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
