

Molecular conflicts disrupting centromere maintenance contribute to Xenopus hybrid inviability
- PMID: 35973429
- PMCID: PMC9529917
- DOI: 10.1016/j.cub.2022.07.037
Molecular conflicts disrupting centromere maintenance contribute to Xenopus hybrid inviability
Abstract
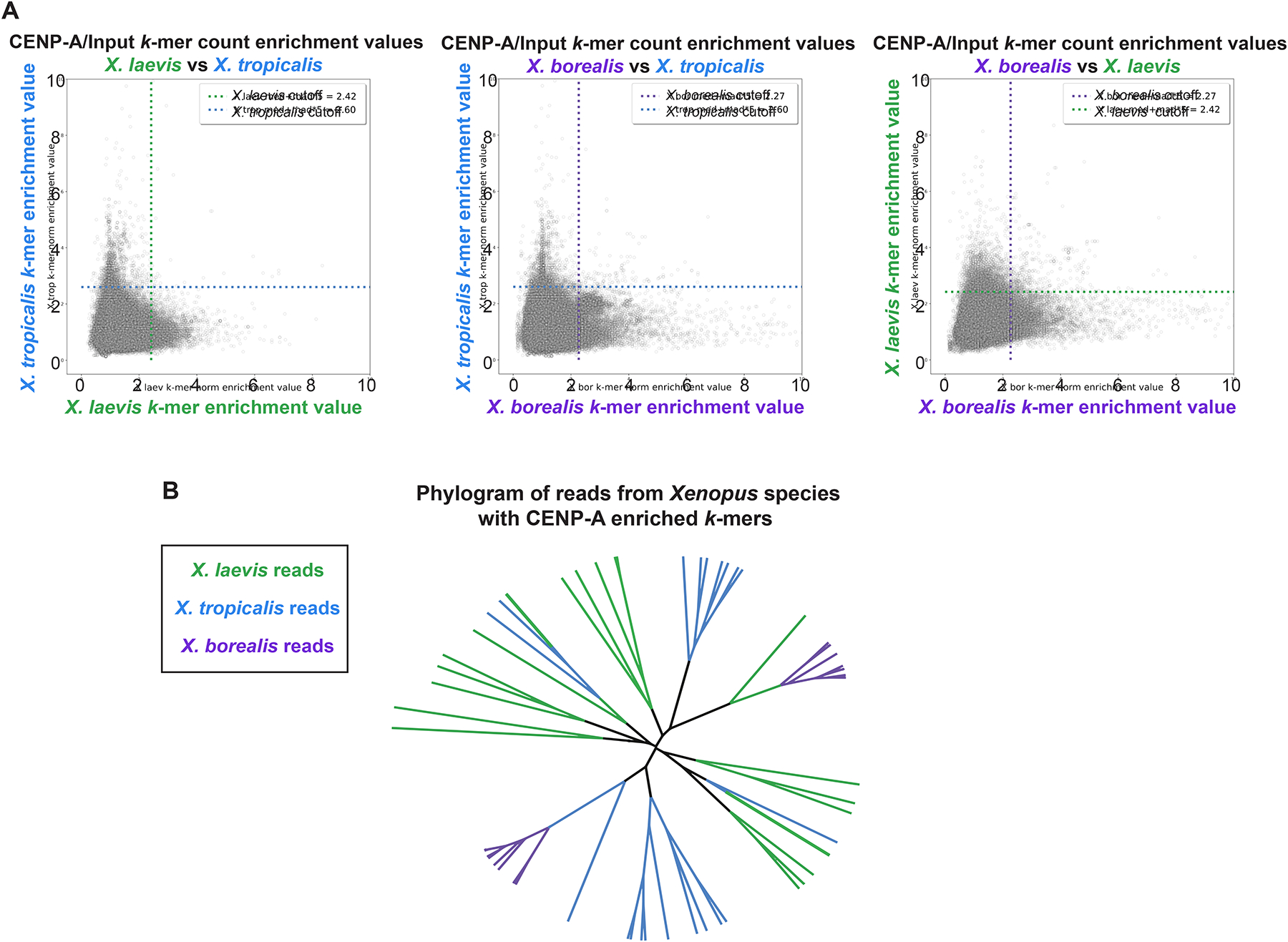
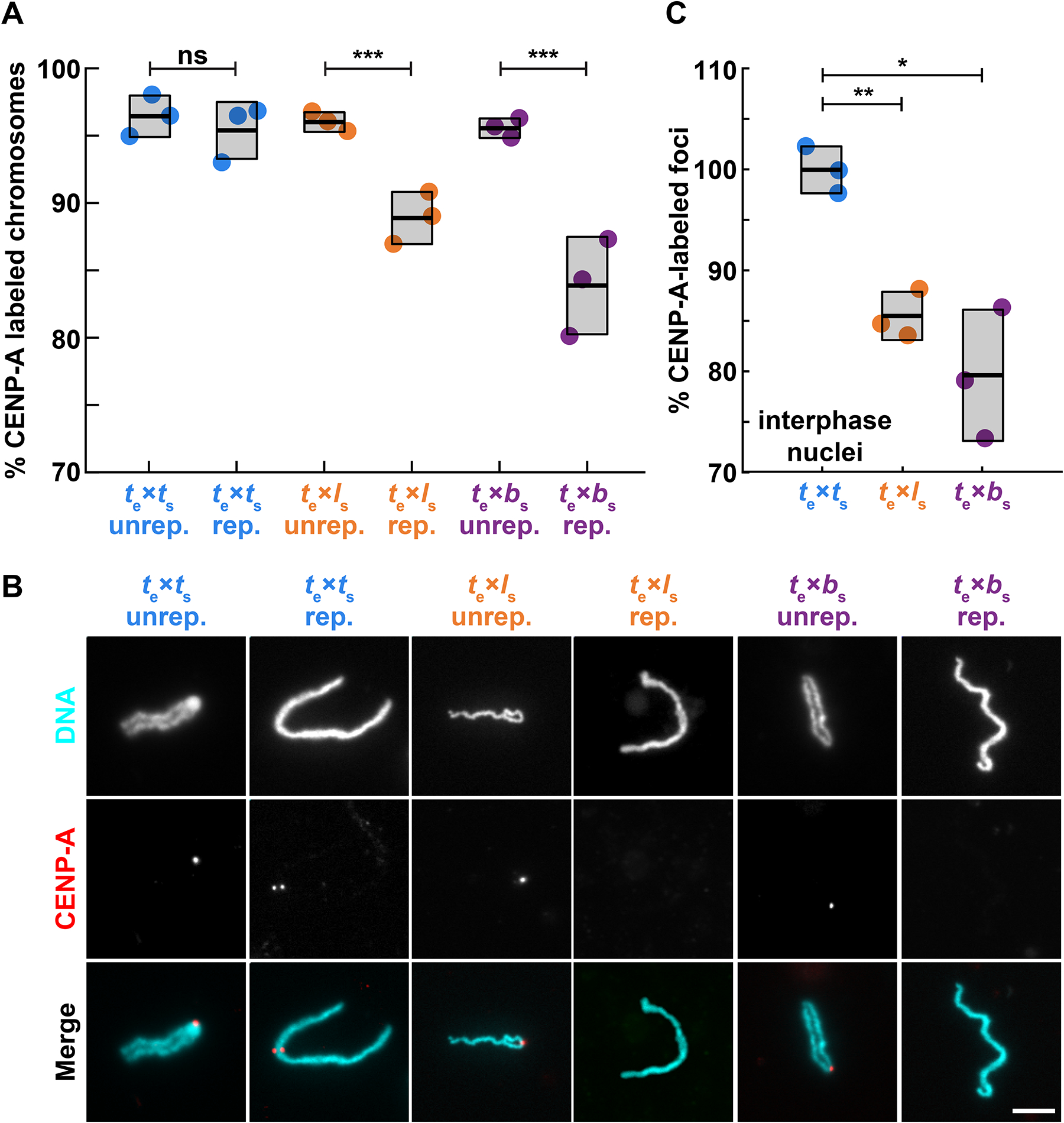
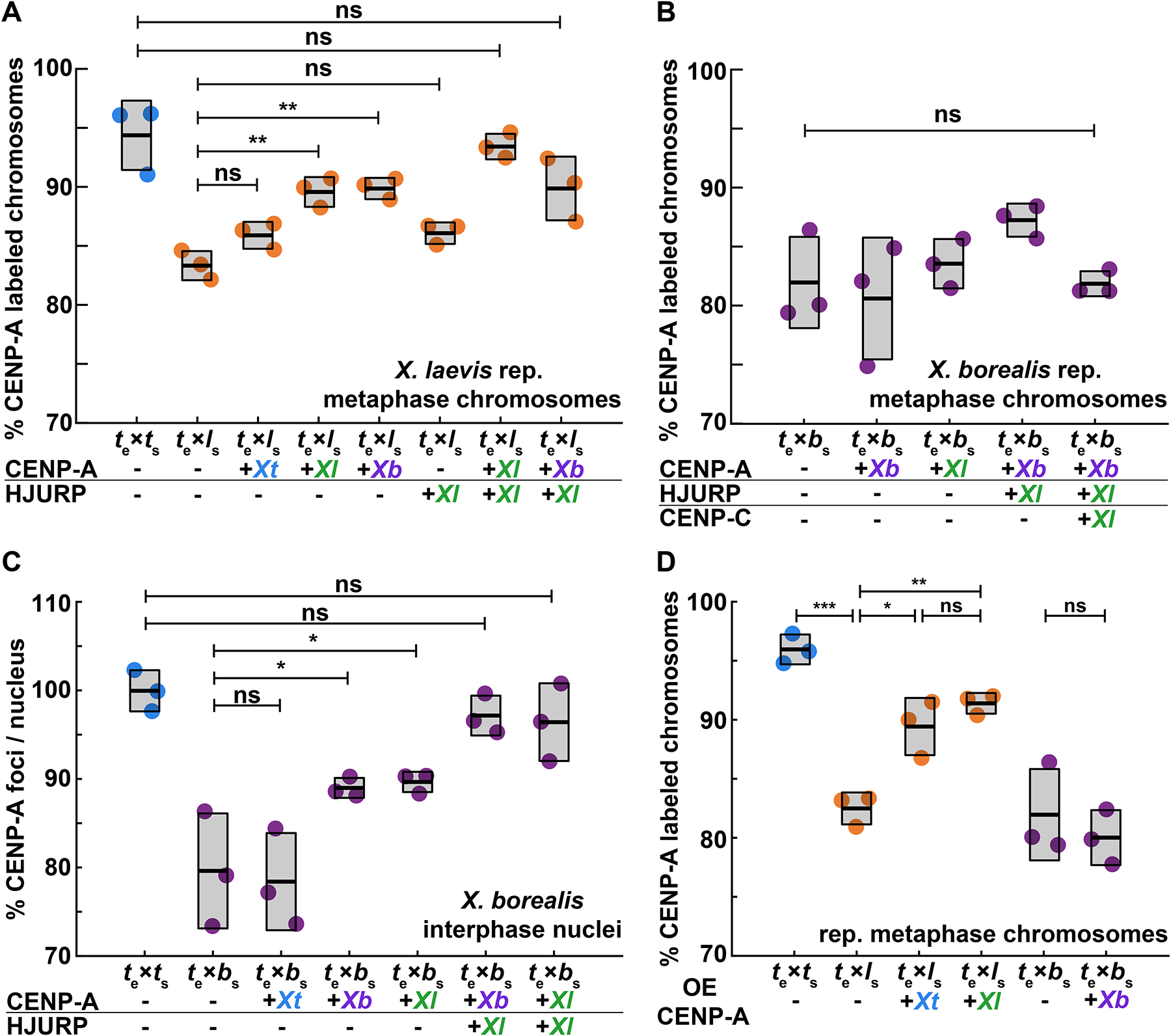
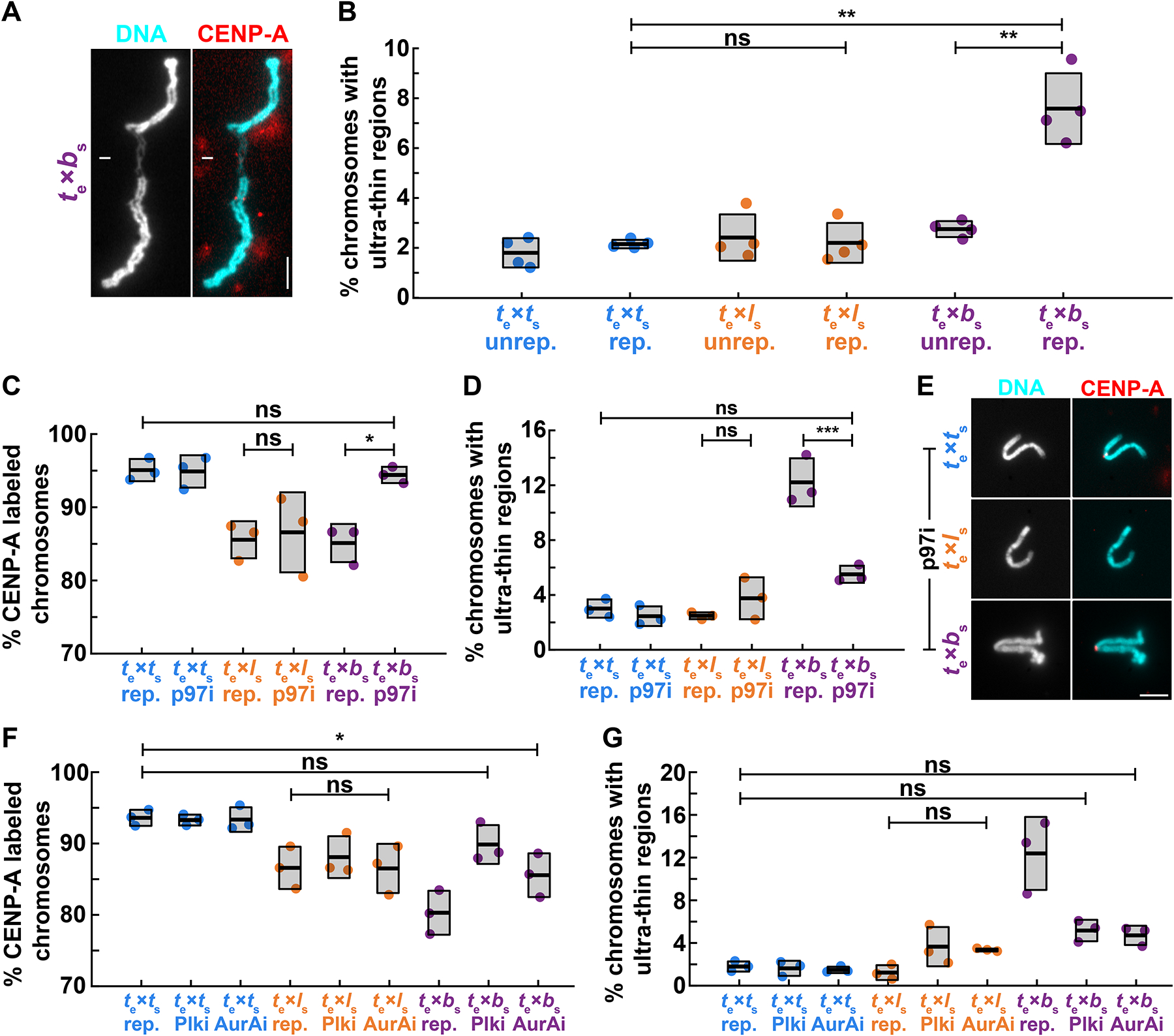
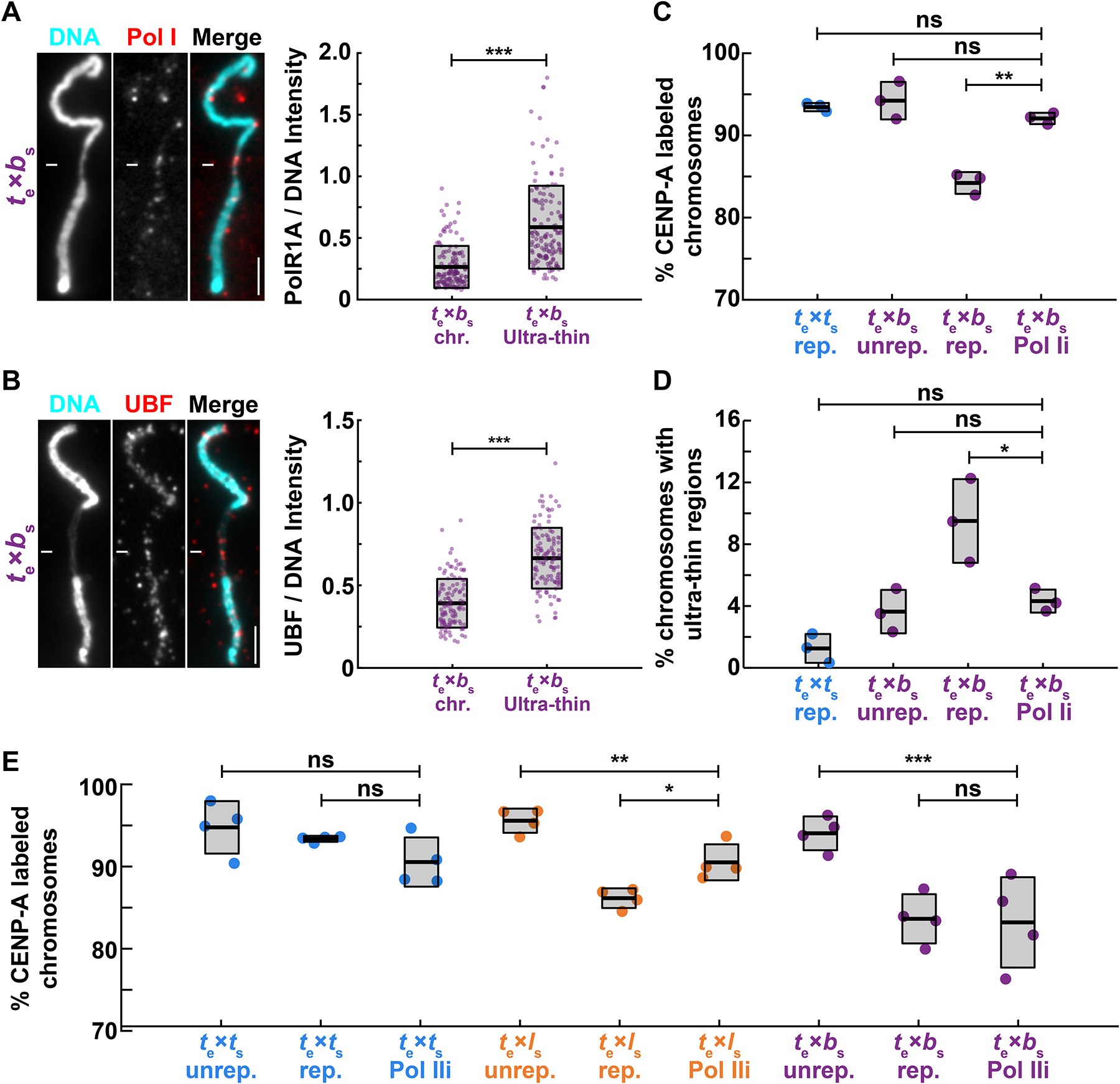
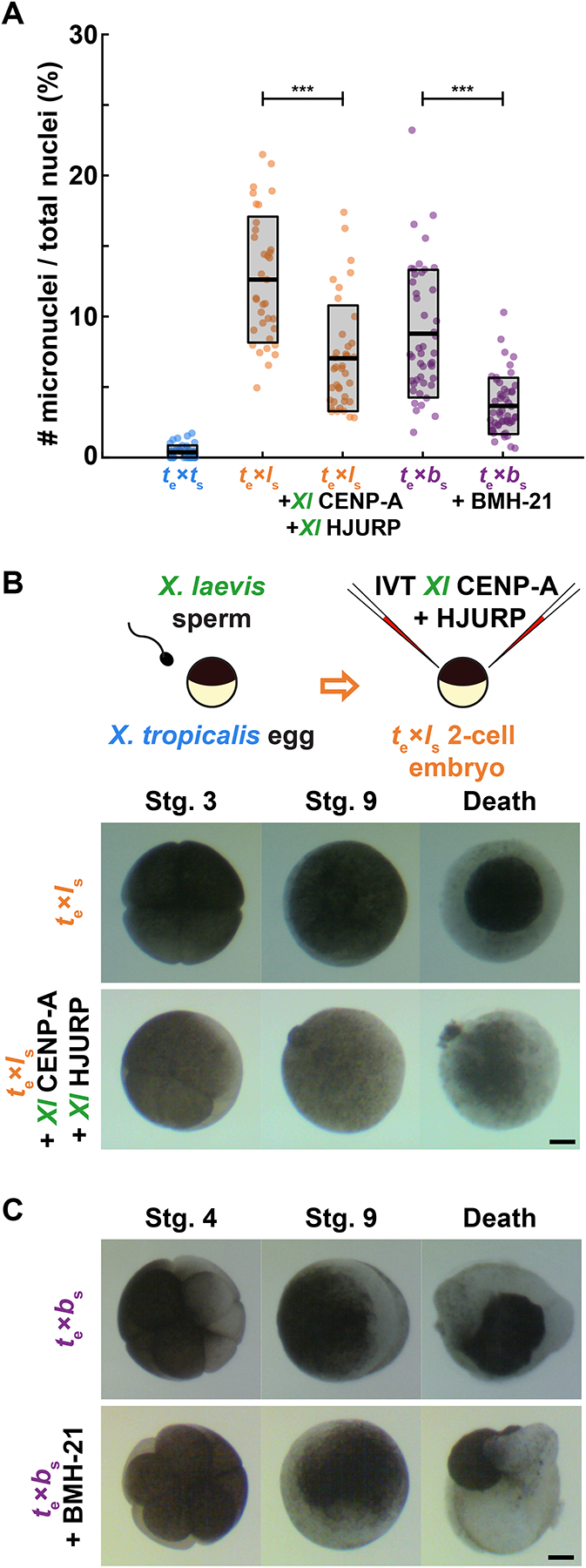
Although central to evolution, the causes of hybrid inviability that drive reproductive isolation are poorly understood. Embryonic lethality occurs when the eggs of the frog X. tropicalis are fertilized with either X. laevis or X. borealis sperm. We observed that distinct subsets of paternal chromosomes failed to assemble functional centromeres, causing their mis-segregation during embryonic cell divisions. Core centromere DNA sequence analysis revealed little conservation among the three species, indicating that epigenetic mechanisms that normally operate to maintain centromere integrity are disrupted on specific paternal chromosomes in hybrids. In vitro reactions combining X. tropicalis egg extract with either X. laevis or X. borealis sperm chromosomes revealed that paternally matched or overexpressed centromeric histone CENP-A and its chaperone HJURP could rescue centromere assembly on affected chromosomes in interphase nuclei. However, although the X. laevis chromosomes maintained centromeric CENP-A in metaphase, X. borealis chromosomes did not and also displayed ultra-thin regions containing ribosomal DNA. Both centromere assembly and morphology of X. borealis mitotic chromosomes could be rescued by inhibiting RNA polymerase I or preventing the collapse of stalled DNA replication forks. These results indicate that specific paternal centromeres are inactivated in hybrids due to the disruption of associated chromatin regions that interfere with CENP-A incorporation, at least in some cases due to conflicts between replication and transcription machineries. Thus, our findings highlight the dynamic nature of centromere maintenance and its susceptibility to disruption in vertebrate interspecies hybrids.
Keywords: CENP-A; Xenopus; centromere; chromosome segregation; hybrid incompatibility; speciation.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Cell biology: Centromere instability and replication stress underlie Xenopus hybrid incompatibility.Curr Biol. 2022 Sep 26;32(18):R947-R949. doi: 10.1016/j.cub.2022.08.015. Curr Biol. 2022. PMID: 36167041
References
-
- Maheshwari S, and Barbash DA (2011). The Genetics of Hybrid Incompatibilities. Annu. Rev. Genet 45, 331–355. - PubMed
-
- Fujiwara A, Abe S, Yamaha E, Yamazaki F, and Yoshida MC (1997). Uniparental chromosome elimination in the early embryogenesis of the inviable salmonid hybrids between masu salmon female and rainbow trout male. Chromosoma 106, 44–52. - PubMed
-
- Gernand D, Rutten T, Varshney A, Rubtsova M, Prodanovic S, and Bru C (2005). Uniparental Chromosome Elimination at Mitosis and Interphase in Wheat and Pearl Millet Crosses Involves Micronucleus Formation, Progressive Heterochromatinization, and DNA Fragmentation. Plant Cell 17, 2431–2438. - PMC - PubMed
