

Copper starvation induces antimicrobial isocyanide integrated into two distinct biosynthetic pathways in fungi
- PMID: 35973982
- PMCID: PMC9381783
- DOI: 10.1038/s41467-022-32394-x
Copper starvation induces antimicrobial isocyanide integrated into two distinct biosynthetic pathways in fungi
Abstract
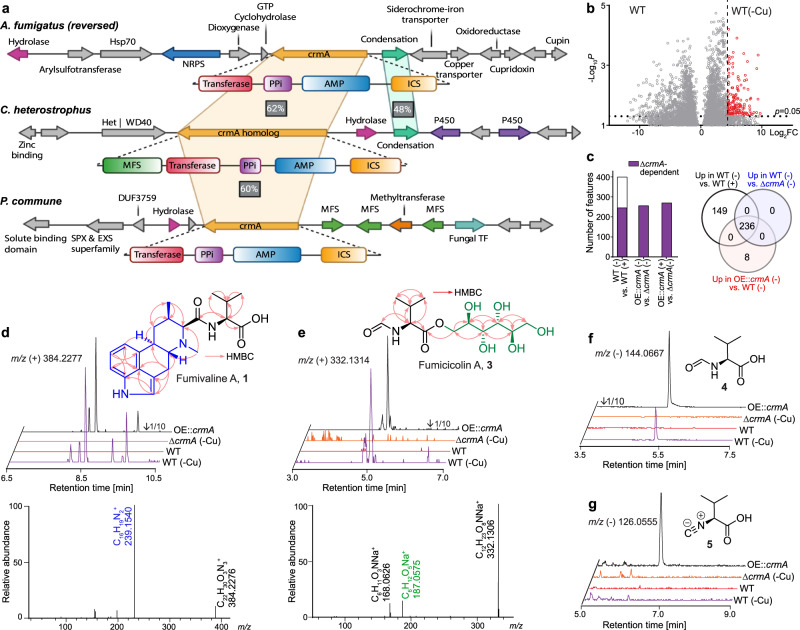
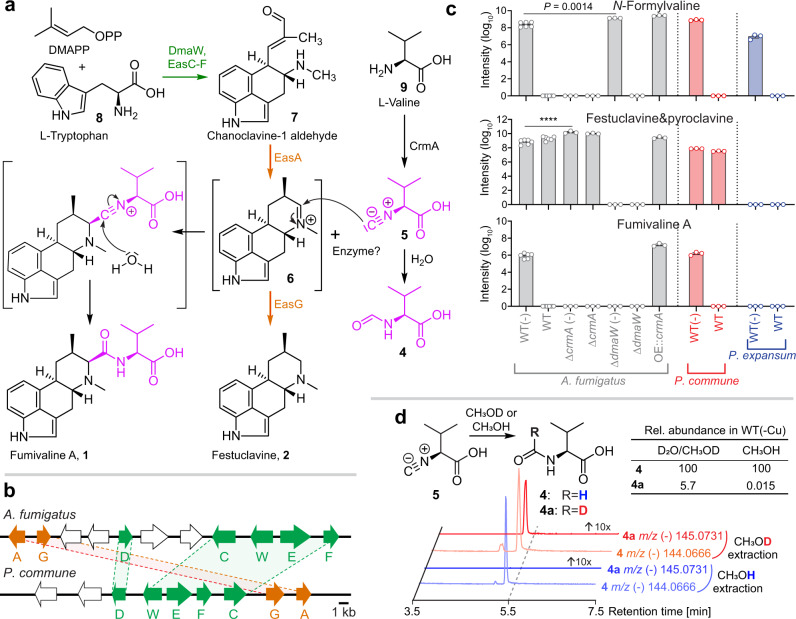
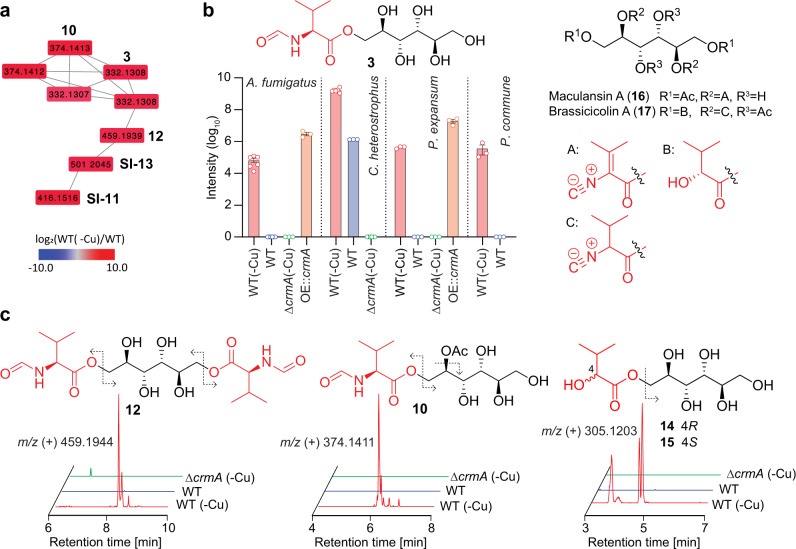
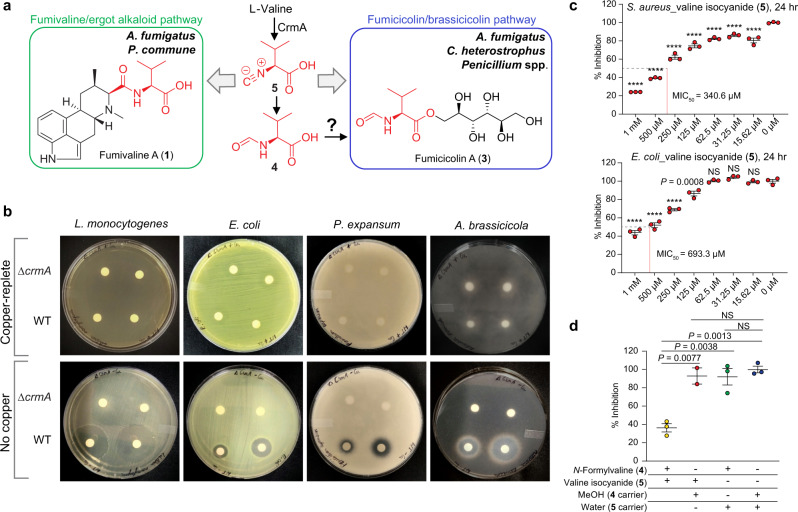
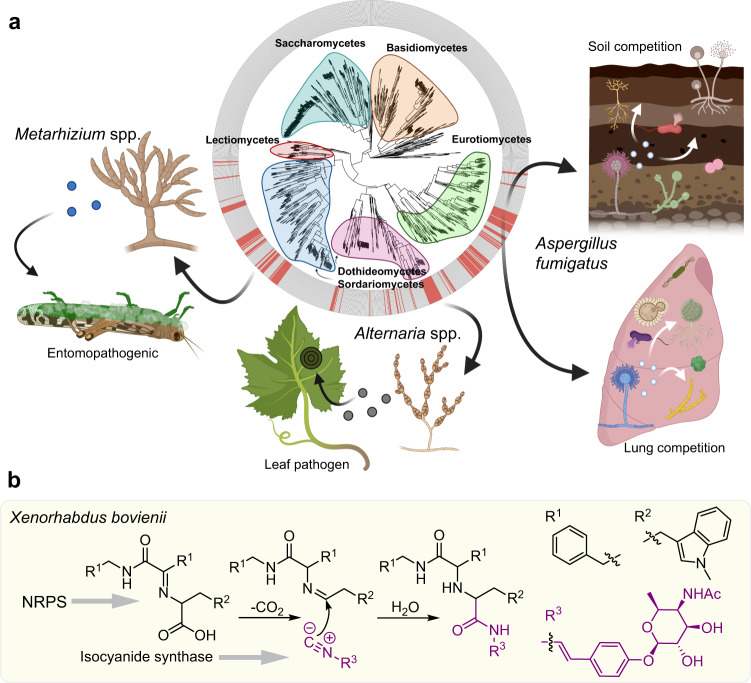
The genomes of many filamentous fungi, such as Aspergillus spp., include diverse biosynthetic gene clusters of unknown function. We previously showed that low copper levels upregulate a gene cluster that includes crmA, encoding a putative isocyanide synthase. Here we show, using untargeted comparative metabolomics, that CrmA generates a valine-derived isocyanide that contributes to two distinct biosynthetic pathways under copper-limiting conditions. Reaction of the isocyanide with an ergot alkaloid precursor results in carbon-carbon bond formation analogous to Strecker amino-acid synthesis, producing a group of alkaloids we term fumivalines. In addition, valine isocyanide contributes to biosynthesis of a family of acylated sugar alcohols, the fumicicolins, which are related to brassicicolin A, a known isocyanide from Alternaria brassicicola. CrmA homologs are found in a wide range of pathogenic and non-pathogenic fungi, some of which produce fumicicolin and fumivaline. Extracts from A. fumigatus wild type (but not crmA-deleted strains), grown under copper starvation, inhibit growth of diverse bacteria and fungi, and synthetic valine isocyanide shows antibacterial activity. CrmA thus contributes to two biosynthetic pathways downstream of trace-metal sensing.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
