

Single cell spatial analysis reveals the topology of immunomodulatory purinergic signaling in glioblastoma
- PMID: 35973991
- PMCID: PMC9381513
- DOI: 10.1038/s41467-022-32430-w
Single cell spatial analysis reveals the topology of immunomodulatory purinergic signaling in glioblastoma
Abstract
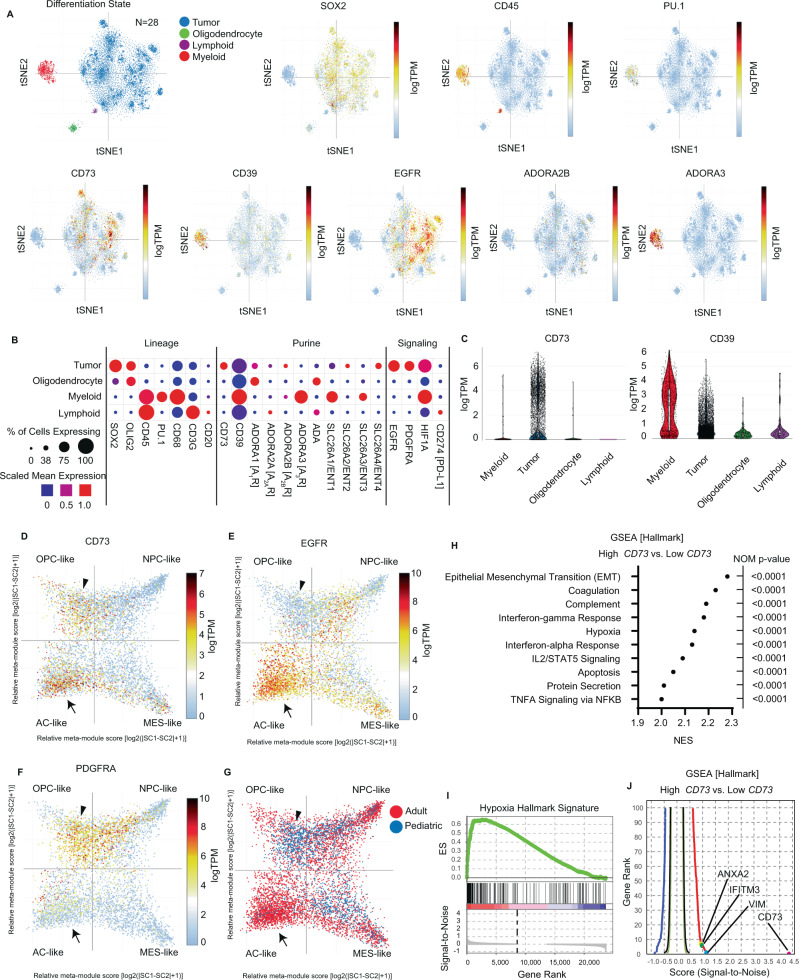
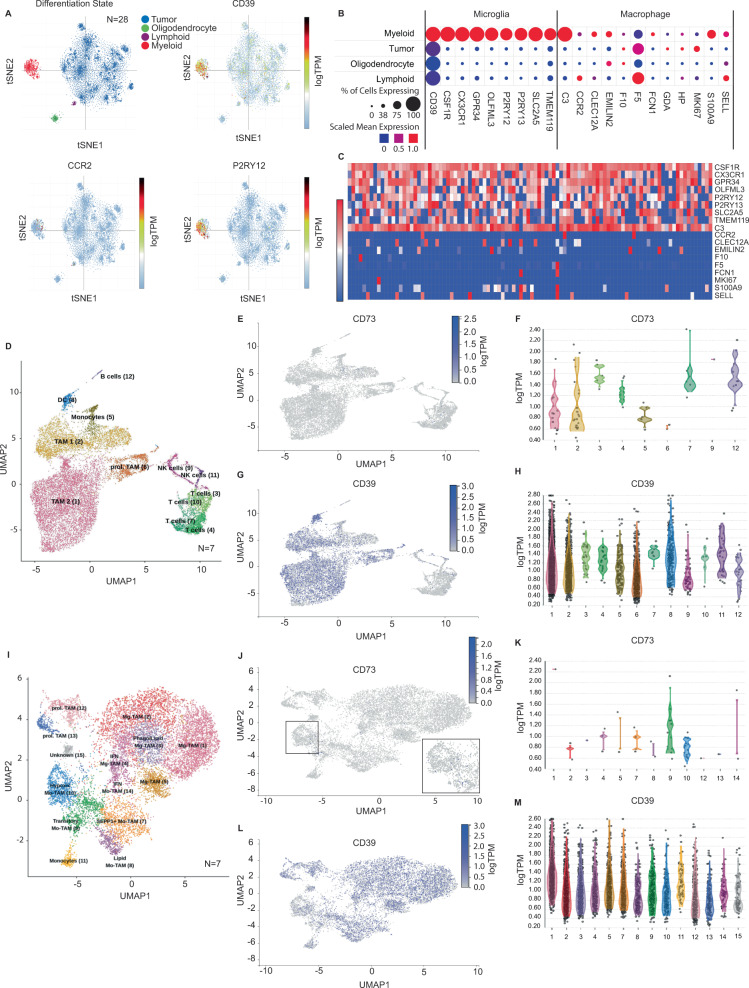
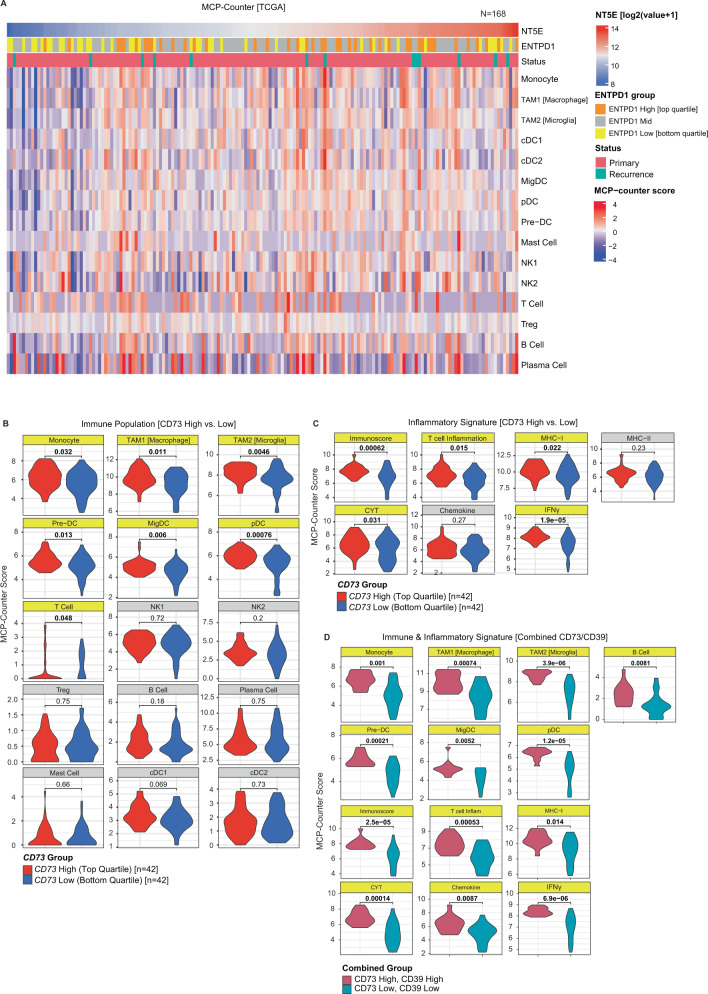
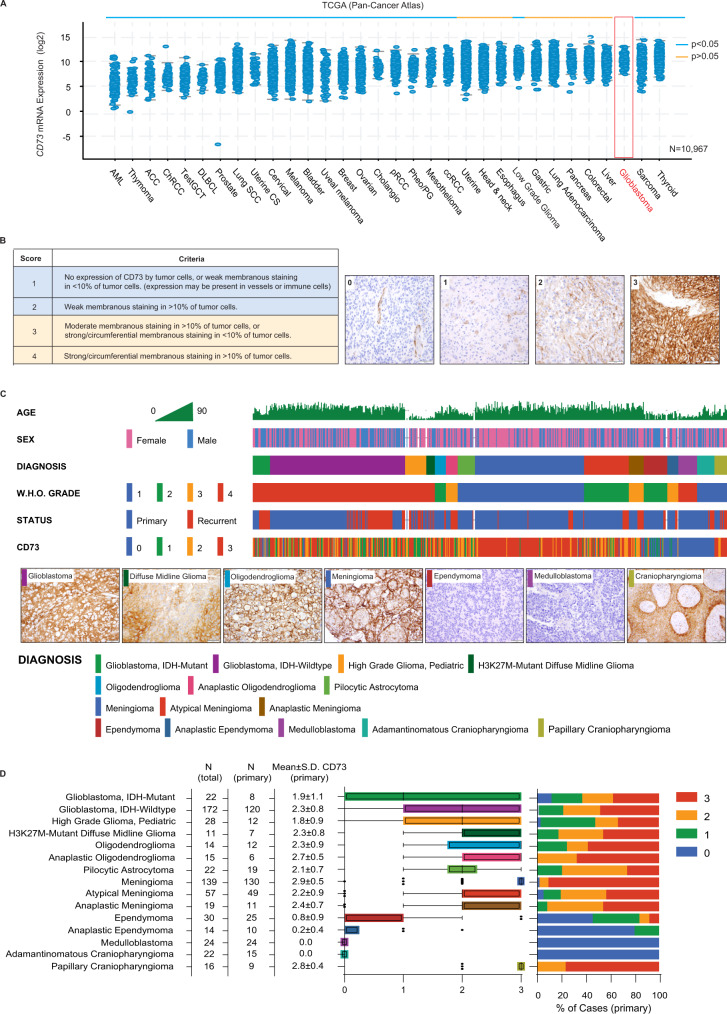
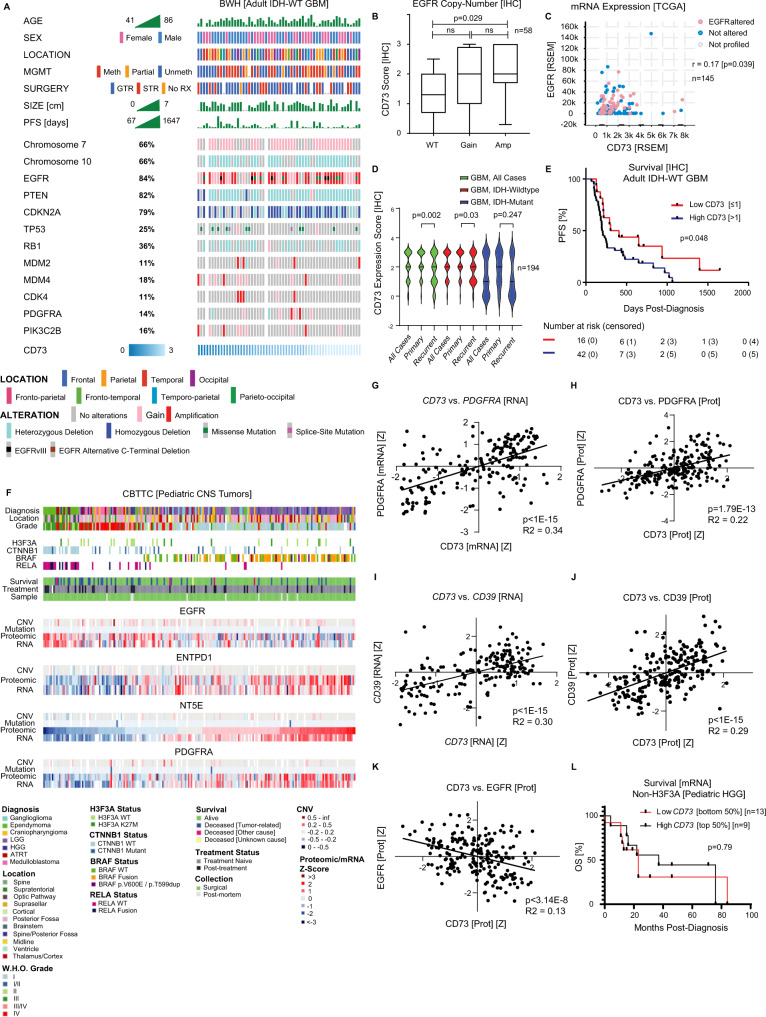
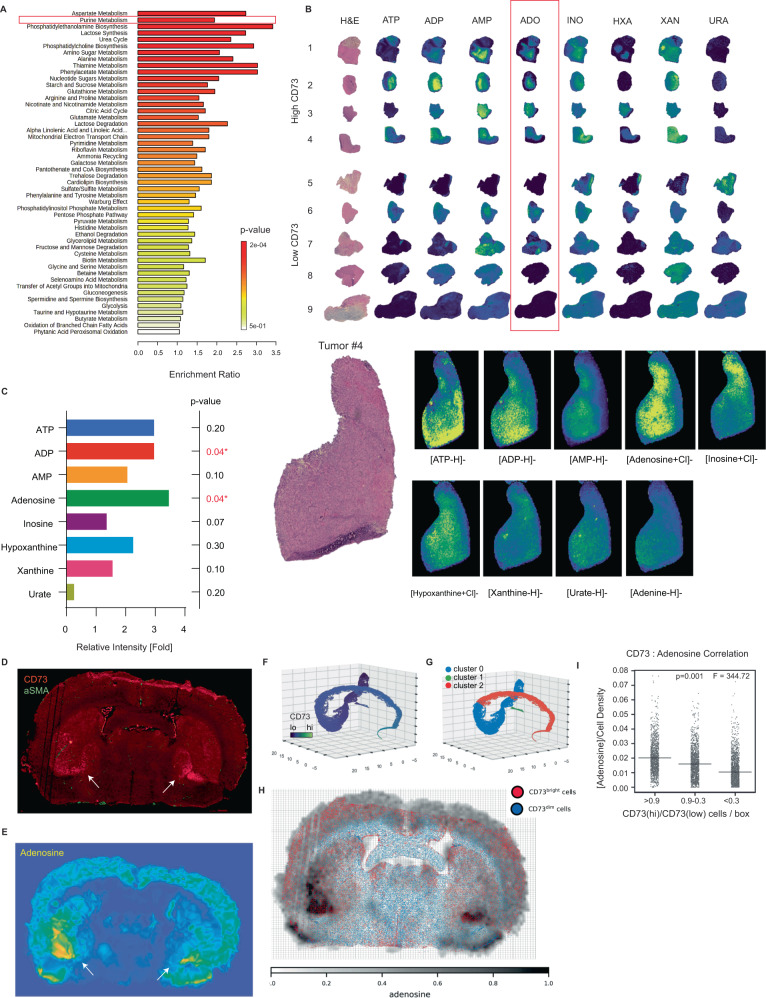
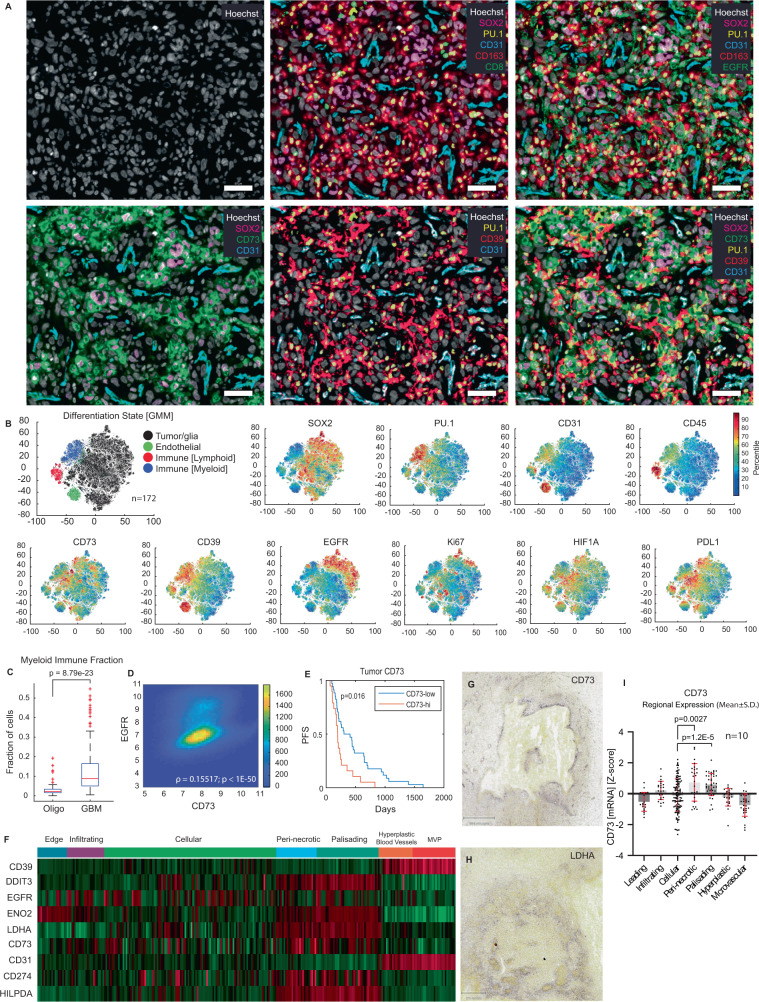
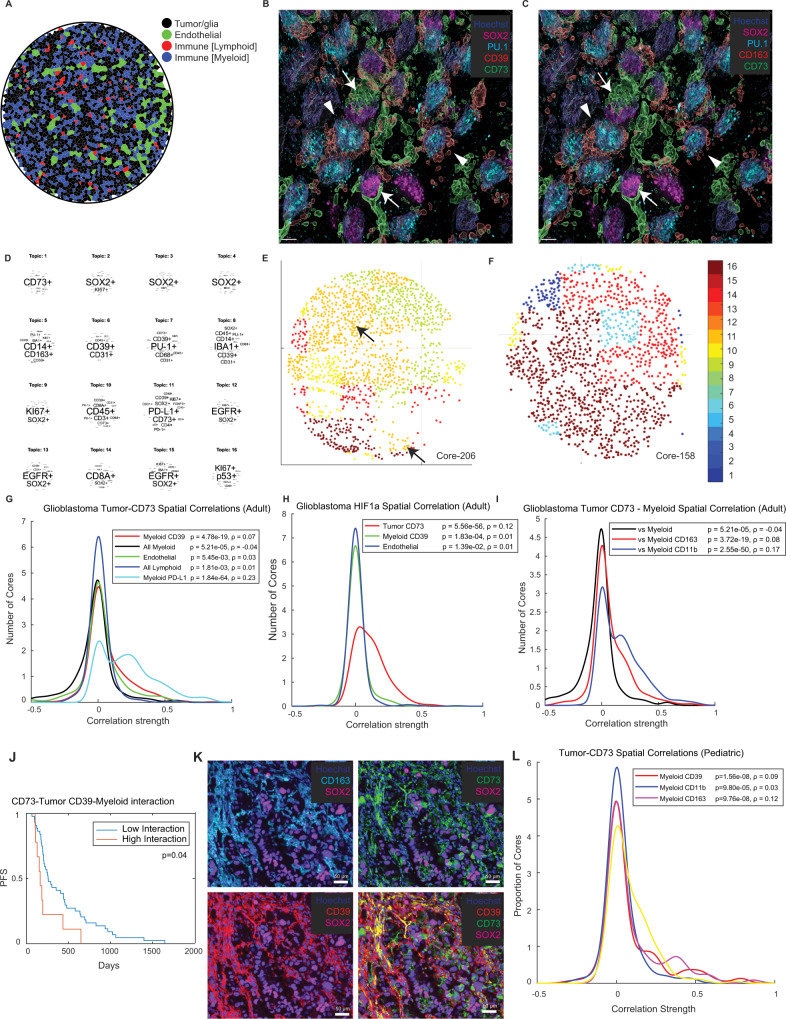
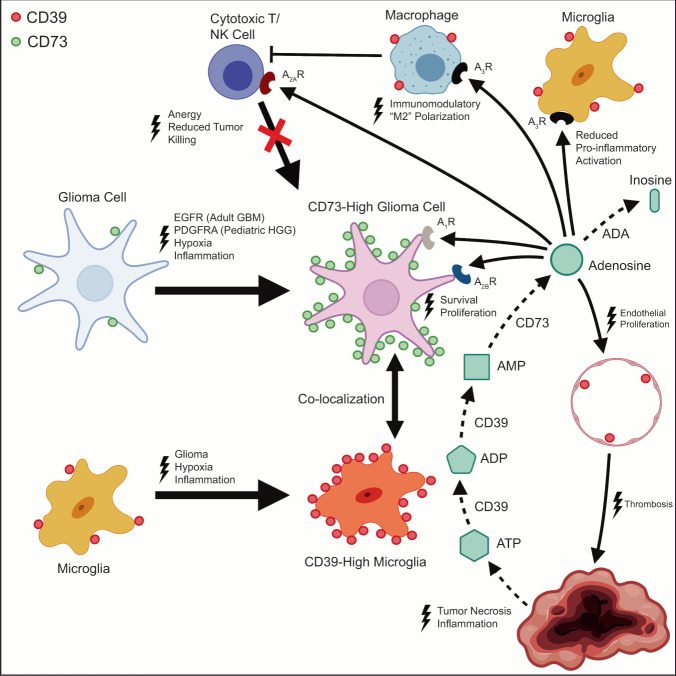
How the glioma immune microenvironment fosters tumorigenesis remains incompletely defined. Here, we use single-cell RNA-sequencing and multiplexed tissue-imaging to characterize the composition, spatial organization, and clinical significance of extracellular purinergic signaling in glioma. We show that microglia are the predominant source of CD39, while tumor cells principally express CD73. In glioblastoma, CD73 is associated with EGFR amplification, astrocyte-like differentiation, and increased adenosine, and is linked to hypoxia. Glioblastomas enriched for CD73 exhibit inflammatory microenvironments, suggesting that purinergic signaling regulates immune adaptation. Spatially-resolved single-cell analyses demonstrate a strong spatial correlation between tumor-CD73 and microglial-CD39, with proximity associated with poor outcomes. Similar spatial organization is present in pediatric high-grade gliomas including H3K27M-mutant diffuse midline glioma. These data reveal that purinergic signaling in gliomas is shaped by genotype, lineage, and functional state, and that core enzymes expressed by tumor and myeloid cells are organized to promote adenosine-rich microenvironments potentially amenable to therapeutic targeting.
© 2022. The Author(s).
Conflict of interest statement
M.T. has a consulting or advisory role with Agios Pharmaceutical, Integragen, and Taiho Oncology, and research funding from Sanofi. P.K.S. is a member of the SAB or BOD member of Applied Biomath, RareCyte Inc., and Glencoe Software; P.K.S. is also a member of the NanoString SAB and a consultant for Montai Health and Merck. In the last five years the Sorger lab has received research funding from Novartis and Merck. KLL is supported by Eli Lilly and BMS, consults for BMS, Integragen, Travera LLC, is on the SAB of Integragen and Rarecyte, and holds equity in Travera LLC. P.B. receives grant funding from Novartis Institute of Biomedical Research and Deerfield, and consults for QED Therapeutics, for unrelated projects. P.Y.W. has research support from Agios, Astra Zeneca, Medimmune, Celgene, Eli Lilly, Genentech, Roche, Kazia, MediciNova, Merck, Novartis, Nuvation Bio, Chimerix, Vascular Biogenics, and VBI Vaccines. P.W. is on the advisory boards of Agios, Astra Zeneca, Black Diamond, Boston Pharmaceuticals, Chimerix, CNS Pharmaceuticals, Elevate Bio, Imvax, Karyopharm, Merck, Mundipharma, Novocure, Novartis, Nuvation Bio, Prelude Therapeutics, Vascular Biogenics, VBI Vaccines, Voyager, and QED. RareCyte manufactured instruments used for tissue imaging and Glencoe developed the OMERO Database used for image informatics. P.K.S. and S.S. declare that none of these relationships have influenced the content of this manuscript. The remaining authors declare no competing interests.
Figures
References
-
- Sharma P, et al. The next decade of immune checkpoint therapy. Cancer Discov. 2021;11:838–857. doi: 10.1158/2159-8290.CD-20-1680. - DOI - PubMed
-
- von Bernstorff W, et al. Systemic and local immunosuppression in pancreatic cancer patients. Clin. Cancer Res. 2001;7:925s–932s. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- T32 CA009216/CA/NCI NIH HHS/United States
- T32 GM008208/GM/NIGMS NIH HHS/United States
- T32 GM008313/GM/NIGMS NIH HHS/United States
- R01 CA194005/CA/NCI NIH HHS/United States
- U2C DK119886/DK/NIDDK NIH HHS/United States
- U54 CA210180/CA/NCI NIH HHS/United States
- T32 EB025823/EB/NIBIB NIH HHS/United States
- U54 CA225088/CA/NCI NIH HHS/United States
- P01 CA163205/CA/NCI NIH HHS/United States
- R01 CA219943/CA/NCI NIH HHS/United States
- P41 EB028741/EB/NIBIB NIH HHS/United States
- P30 CA006516/CA/NCI NIH HHS/United States
- T32 GM007748/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
