

Flight or protection: the genes Ultrabithorax and apterous in the determination of membranous and sclerotized wings in insects
- PMID: 35975435
- PMCID: PMC9382207
- DOI: 10.1098/rspb.2022.0967
Flight or protection: the genes Ultrabithorax and apterous in the determination of membranous and sclerotized wings in insects
Abstract
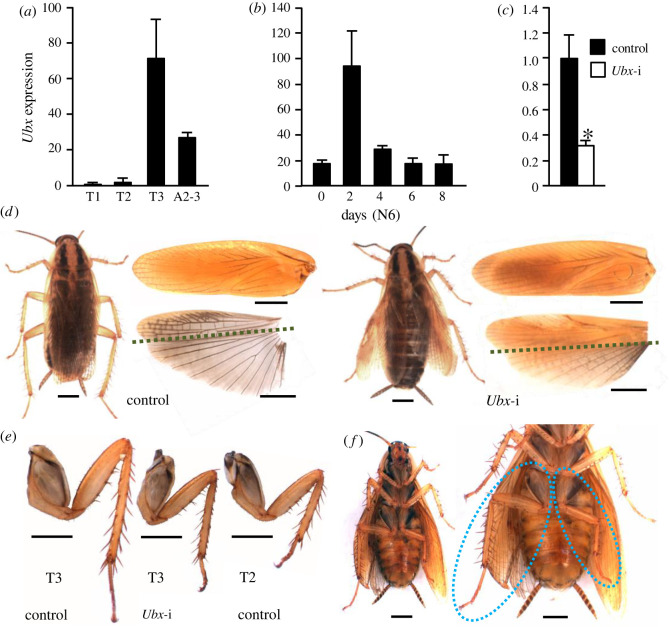
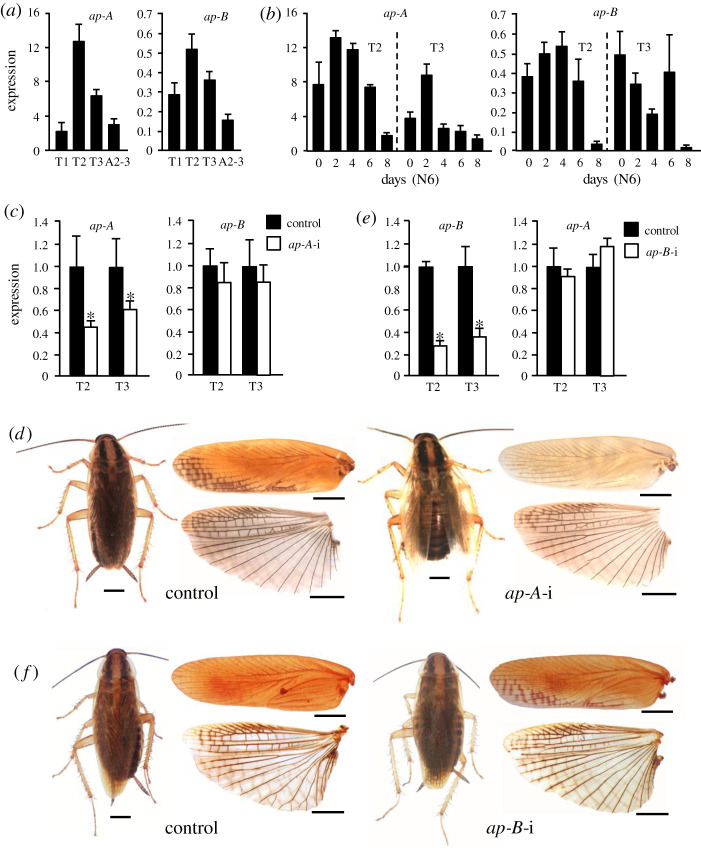
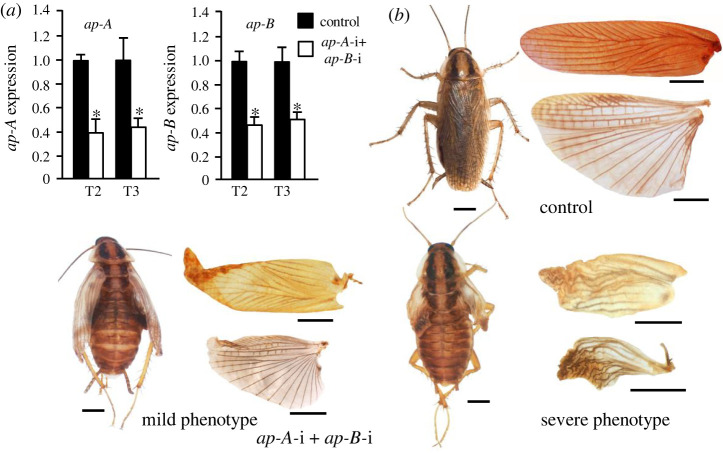
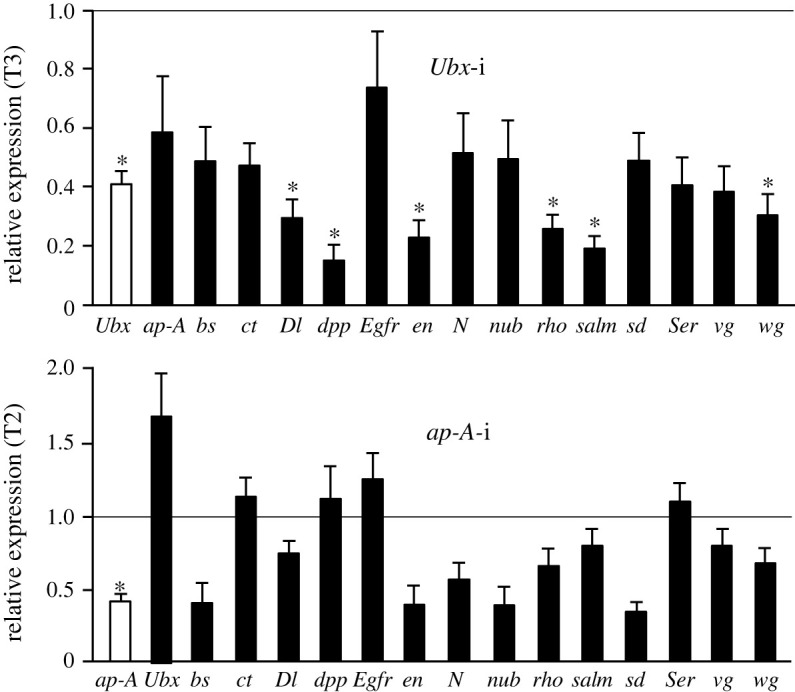
Present-day pterygote insects have two pairs of wings, one in the mesothorax (T2), the other in the metathorax (T3), and both have diverged in structure and function in different groups. Studies in endopterygote and paraneopteran species have shown that the gene Ultrabithorax (Ubx) specifies the identity and wing structure in T3, whereas the gene apterous (ap) significantly contributes to forming modified T2 wings. We wondered whether these Ubx and ap mechanisms operate in the lineage of polyneopterans. To explore this possibility, we used the cockroach Blattella germanica (Polyneoptera and Blattodea), in which the T2 wings are sclerotized (tegmina), whereas those of the T3 are membranous. We found that Ubx determines the structure of T3 and the membranous wing, while ap significantly contributes to form the sclerotized T2 tegmina. These results along with the studies carried out on the beetle Tribolium castaneum by Tomoyasu and collaborators suggest that ap plays an important role in the sclerotization and melanization of the T2 wings in neopteran groups that have sclerotized forewings. In turn, the sclerotizing properties of ap demonstrated in beetles and cockroaches suggest that the origin of this function goes back to the emergence of Neoptera, in the mid Devonian.
Keywords: Apterous; Blattella; Ultrabithorax; insect wings; tribolium; wing specialization.
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Ultrabithorax is required for membranous wing identity in the beetle Tribolium castaneum.Nature. 2005 Feb 10;433(7026):643-7. doi: 10.1038/nature03272. Nature. 2005. PMID: 15703749
-
Tergal and pleural structures contribute to the formation of ectopic prothoracic wings in cockroaches.R Soc Open Sci. 2016 Aug 3;3(8):160347. doi: 10.1098/rsos.160347. eCollection 2016 Aug. R Soc Open Sci. 2016. PMID: 27853616 Free PMC article.
-
Tergal and pleural wing-related tissues in the German cockroach and their implication to the evolutionary origin of insect wings.Evol Dev. 2021 Mar;23(2):100-116. doi: 10.1111/ede.12372. Epub 2021 Jan 27. Evol Dev. 2021. PMID: 33503322
-
Hox and wings.Bioessays. 2005 Jul;27(7):673-5. doi: 10.1002/bies.20260. Bioessays. 2005. PMID: 15954097 Review.
-
Ultrabithorax and the evolution of insect forewing/hindwing differentiation.Curr Opin Insect Sci. 2017 Feb;19:8-15. doi: 10.1016/j.cois.2016.10.007. Epub 2016 Nov 2. Curr Opin Insect Sci. 2017. PMID: 28521947 Review.
Cited by
-
300 Million years of coral treaders (Insecta: Heteroptera: Hermatobatidae) back to the ocean in the phylogenetic context of Arthropoda.Proc Biol Sci. 2023 Jun 28;290(2001):20230855. doi: 10.1098/rspb.2023.0855. Epub 2023 Jun 28. Proc Biol Sci. 2023. PMID: 37357866 Free PMC article.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
