

Cascading effects of prey identity on gene expression in a kleptoplastidic ciliate
- PMID: 35975609
- PMCID: PMC10087830
- DOI: 10.1111/jeu.12940
Cascading effects of prey identity on gene expression in a kleptoplastidic ciliate
Erratum in
-
Correction to "Cascading effects of prey identity on gene expression in a kleptoplastidic ciliate".J Eukaryot Microbiol. 2023 May-Jun;70(3):e12968. doi: 10.1111/jeu.12968. Epub 2023 Mar 12. J Eukaryot Microbiol. 2023. PMID: 36908081 Free PMC article. No abstract available.
Abstract
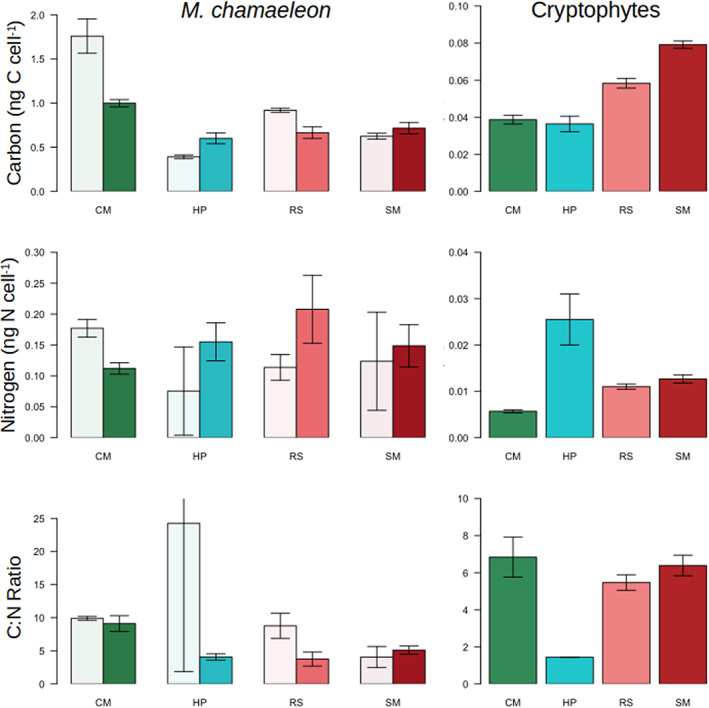
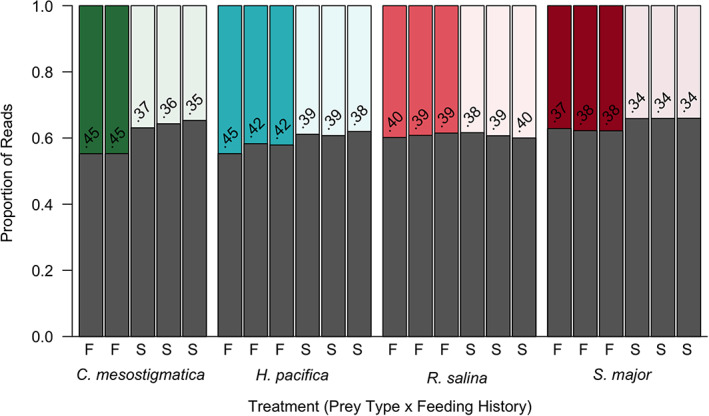
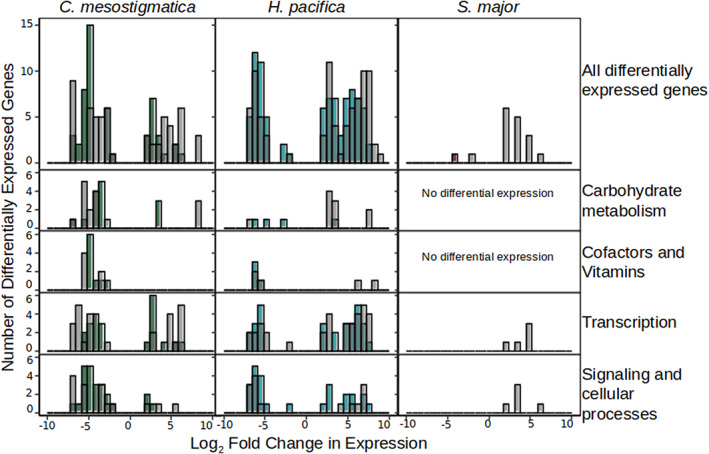
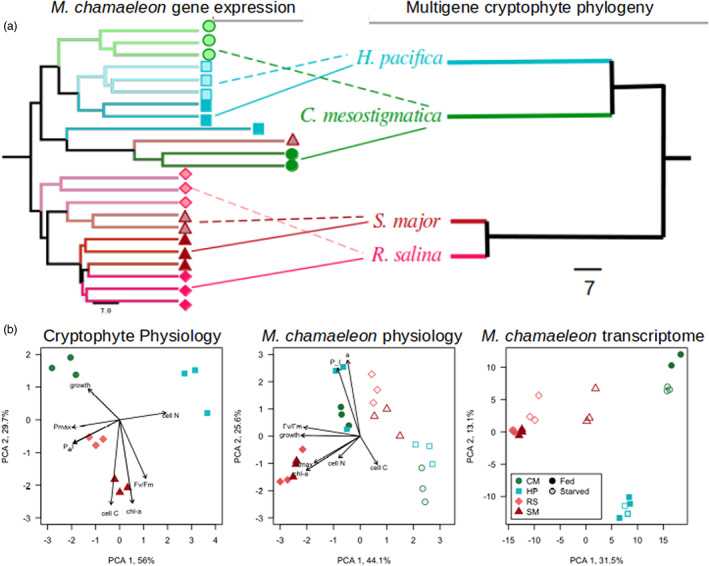
Kleptoplastidic, or chloroplast stealing, lineages transiently retain functional photosynthetic machinery from algal prey. This machinery, and its photosynthetic outputs, must be integrated into the host's metabolism, but the details of this integration are poorly understood. Here, we study this metabolic integration in the ciliate Mesodinium chamaeleon, a coastal marine species capable of retaining chloroplasts from at least six distinct genera of cryptophyte algae. To assess the effects of feeding history on ciliate physiology and gene expression, we acclimated M. chamaeleon to four different types of prey and contrasted well-fed and starved treatments. Consistent with previous physiological work on the ciliate, we found that starved ciliates had lower chlorophyll content, photosynthetic rates, and growth rates than their well-fed counterparts. However, ciliate gene expression mirrored prey phylogenetic relationships rather than physiological status, suggesting that, even as M. chamaeleon cells were starved of prey, their overarching regulatory systems remained tuned to the prey type to which they had been acclimated. Collectively, our results indicate a surprising degree of prey-specific host transcriptional adjustments, implying varied integration of prey metabolic potential into many aspects of ciliate physiology.
Keywords: Mesodinium chamaeleon; acquired metabolism; cryptophyte; photophysiology; transcriptomics.
© 2022 The Authors. Journal of Eukaryotic Microbiology published by Wiley Periodicals LLC on behalf of International Society of Protistologists.
Figures
References
-
- Bernhard, J.M. & Bowser, S.S. (1999) Benthic foraminifera of dysoxic sediments: chloroplast sequestration and functional morphology. Earth‐Science Reviews, 46(1), 149–165. 10.1016/S0012-8252(99)00017-3 - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
