

Circadian disruption alters gut barrier integrity via a ß-catenin-MMP-related pathway
- PMID: 35976519
- PMCID: PMC9938043
- DOI: 10.1007/s11010-022-04536-8
Circadian disruption alters gut barrier integrity via a ß-catenin-MMP-related pathway
Abstract
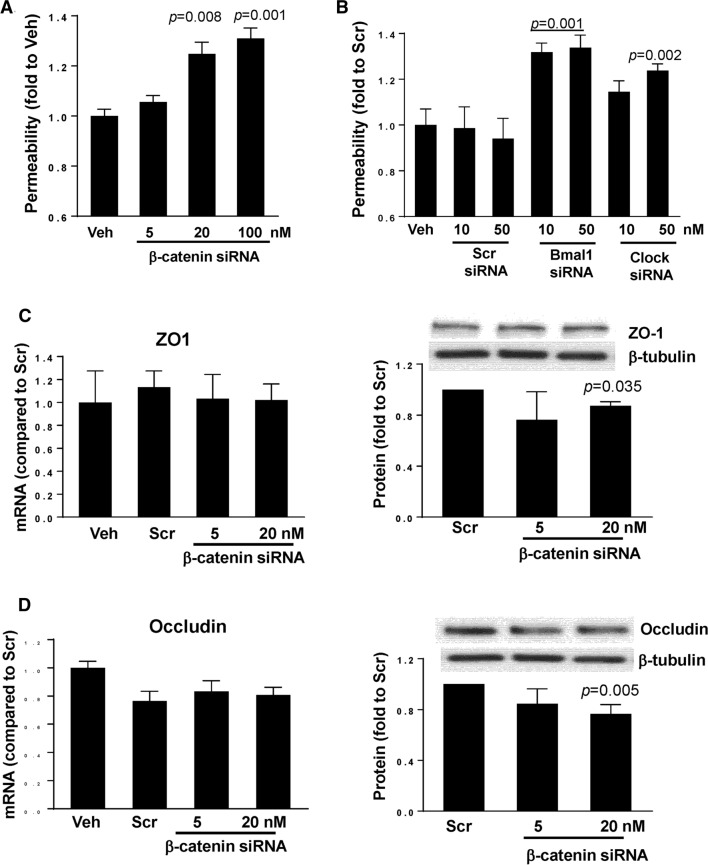
We evaluated the mechanistic link between circadian rhythms and gut barrier permeability. Mice were subjected to either constant 24-h light (LL) or 12-h light/dark cycles (LD). Mice housed in LL experienced a significant increase in gut barrier permeability that was associated with dysregulated ß-catenin expression and altered expression of tight junction (TJ) proteins. Silencing of ß-catenin resulted in disruption of barrier function in SW480 cells, with ß-catenin appearing to be an upstream regulator of the core circadian components, such as Bmal1, Clock, and Per1/2. In addition, ß-catenin silencing downregulated ZO-1 and occludin TJ proteins with only limited or no changes at their mRNA levels, suggesting post transcriptional regulation. Indeed, silencing of ß-catenin significantly upregulated expression of matrix metallopeptidase (MMP)-2 and MMP-9, and blocking MMP-2/9 activity attenuated epithelial disruption induced by ß-catenin silencing. These results indicate the regulatory role of circadian disruption on gut barrier integrity and the associations between TJ proteins and circadian rhythms, while demonstrating the regulatory role of ß-catenin in this process.
Keywords: Circadian clock genes; Circadian rhythm disruption; Circadian rhythm molecules; Intestinal barrier integrity; Tight junction proteins; ß-Catenin.
© 2022. The Author(s).
Conflict of interest statement
The authors report no competing interests.
Figures







References
-
- Konturek PC, Brzozowski T, Konturek SJ. Gut clock: Implication of circadian rhythms in the gastointestinal tract. J Physiol Pharmacol. 2011;62:39–50. - PubMed
MeSH terms
Substances
Grants and funding
- R01 MH128022/MH/NIMH NIH HHS/United States
- DA050528/DA/NIDA NIH HHS/United States
- DA047157/DA/NIDA NIH HHS/United States
- MH128022/MH/NIMH NIH HHS/United States
- R01 DA039576/DA/NIDA NIH HHS/United States
- R01 DA050528/DA/NIDA NIH HHS/United States
- R01 DA040537/DA/NIDA NIH HHS/United States
- DA044579/DA/NIDA NIH HHS/United States
- MH072567/MH/NIMH NIH HHS/United States
- R01 DA044579/DA/NIDA NIH HHS/United States
- DA039576/DA/NIDA NIH HHS/United States
- MH122235/MH/NIMH NIH HHS/United States
- R01 HL126559/HL/NHLBI NIH HHS/United States
- DA040537/DA/NIDA NIH HHS/United States
- R21 MH122235/MH/NIMH NIH HHS/United States
- R01 DA047157/DA/NIDA NIH HHS/United States
- HL126559/HL/NHLBI NIH HHS/United States
- R01 MH072567/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
