

Global crotonylome reveals hypoxia-mediated lamin A crotonylation regulated by HDAC6 in liver cancer
- PMID: 35977926
- PMCID: PMC9385620
- DOI: 10.1038/s41419-022-05165-1
Global crotonylome reveals hypoxia-mediated lamin A crotonylation regulated by HDAC6 in liver cancer
Abstract
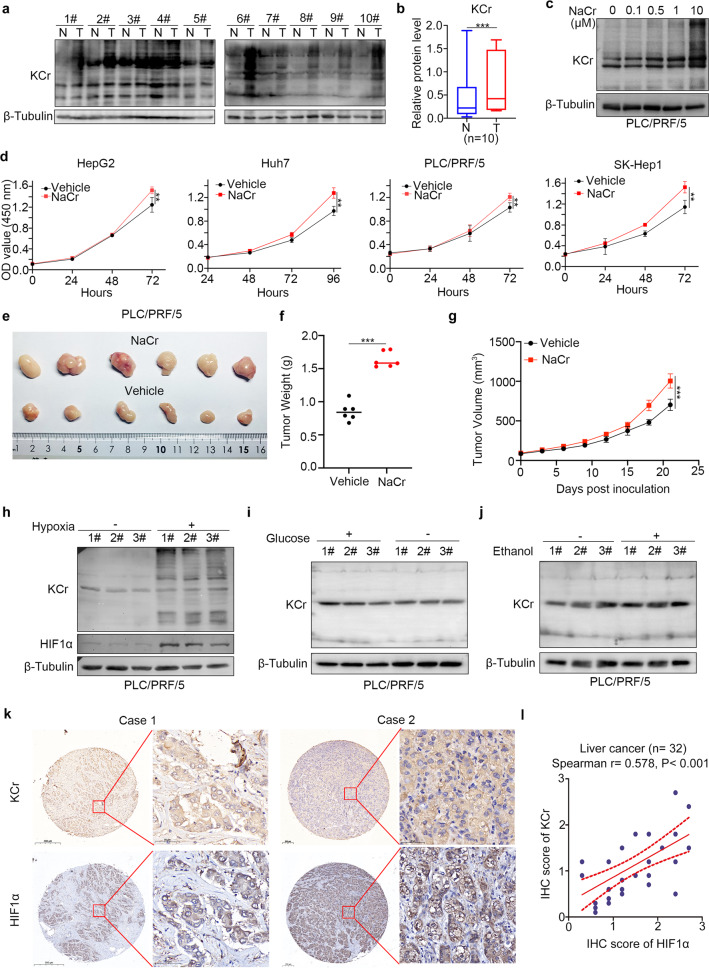
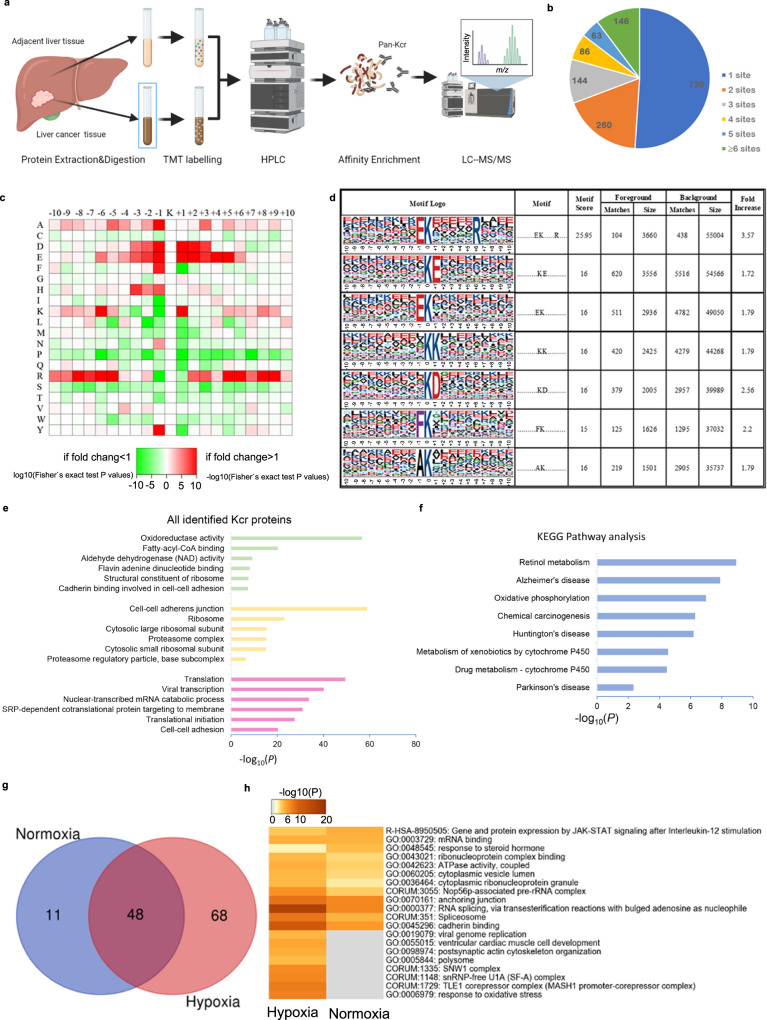
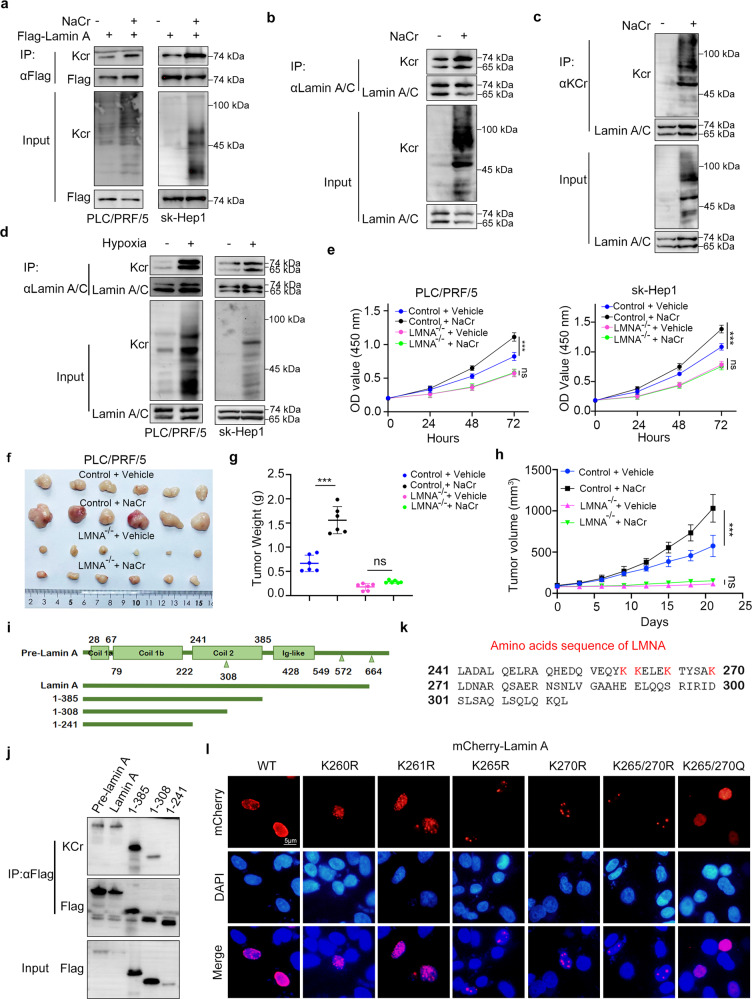
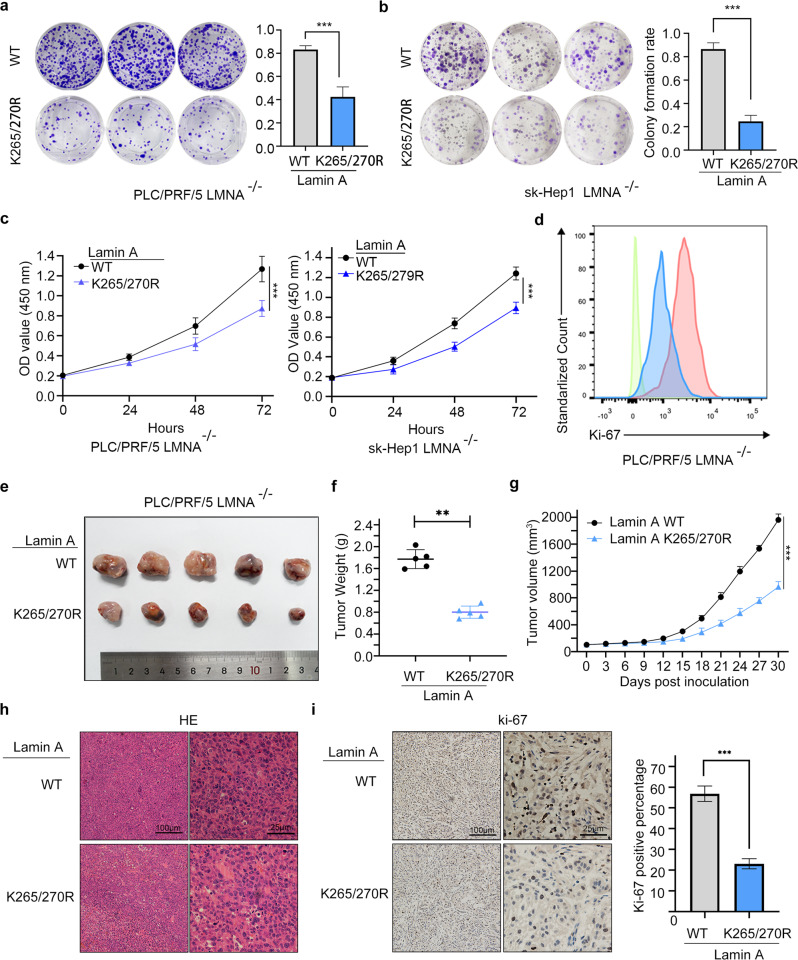
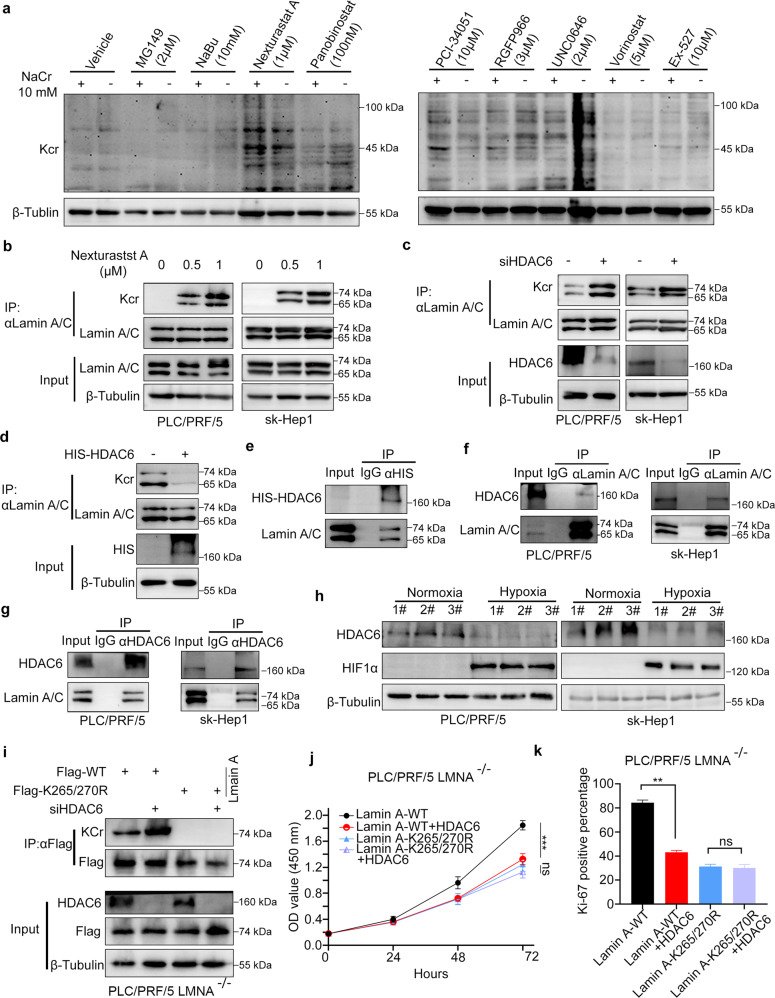
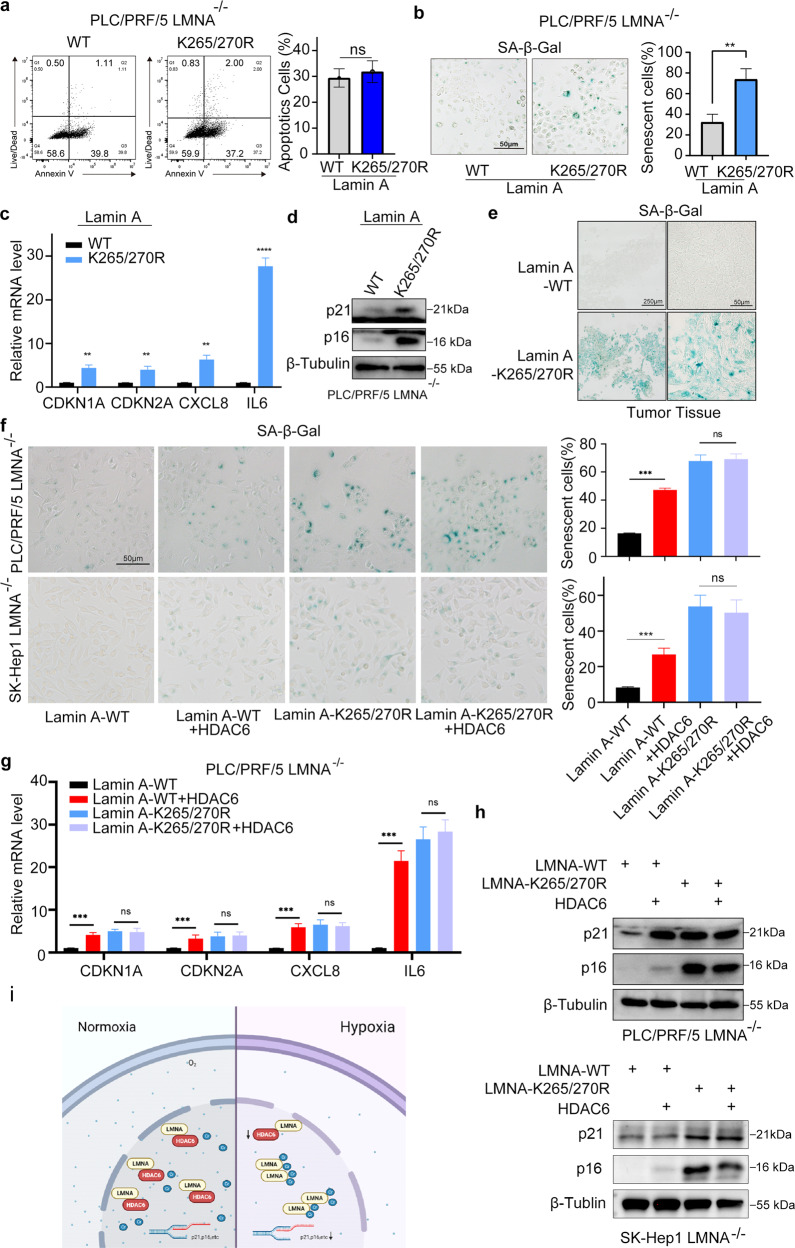
Lysine crotonylation is a recently discovered post-translation modification involved in transcription regulation, cell signal transduction, and other processes. Scientists have identified several crotonylases and decrotonylases of histones, including P300/CBP, HDACs, and SIRTs. However, the regulation of non-histone protein crotonylation remains unclear. In the current study, we verified that crotonylation was upregulated in hypoxia and promoted liver cancer cell growth. We performed TMT-labeled quantitative lysine crotonylome analysis in 12 pairs of hepatocellular carcinoma and adjacent liver tissue and identified 3,793 lysine crotonylation sites in 1,428 proteins. We showed that crotonylation of lamin A at the site of K265/270 maintains its subcellular position, promotes liver cancer cell proliferation, and prevents cellular senescence. Our data indicate that HDAC6 is the decrotonylase of lamin A and downregulated in response to hypoxia, resulting in lamin A K265/270cr. Taken together, our study reveals the lamin A crotonylation in liver cancer progression and fills the research gap in non-histone protein crotonylation function.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Ultradeep Lysine Crotonylome Reveals the Crotonylation Enhancement on Both Histones and Nonhistone Proteins by SAHA Treatment.J Proteome Res. 2017 Oct 6;16(10):3664-3671. doi: 10.1021/acs.jproteome.7b00380. Epub 2017 Sep 19. J Proteome Res. 2017. PMID: 28882038
-
Quantitative Crotonylome Analysis Expands the Roles of p300 in the Regulation of Lysine Crotonylation Pathway.Proteomics. 2018 Aug;18(15):e1700230. doi: 10.1002/pmic.201700230. Epub 2018 Jul 11. Proteomics. 2018. PMID: 29932303 Free PMC article.
-
Lysine crotonylation is involved in hepatocellular carcinoma progression.Biomed Pharmacother. 2019 Mar;111:976-982. doi: 10.1016/j.biopha.2018.12.148. Epub 2019 Jan 9. Biomed Pharmacother. 2019. PMID: 30841477
-
Histone crotonylation-centric gene regulation.Epigenetics Chromatin. 2021 Feb 6;14(1):10. doi: 10.1186/s13072-021-00385-9. Epigenetics Chromatin. 2021. PMID: 33549150 Free PMC article. Review.
-
Crotonylation and disease: Current progress and future perspectives.Biomed Pharmacother. 2023 Sep;165:115108. doi: 10.1016/j.biopha.2023.115108. Epub 2023 Jun 29. Biomed Pharmacother. 2023. PMID: 37392654 Review.
Cited by
-
Mechanism-Based Inhibition of Histone Deacetylase 6 by a Selenocyanate Is Subject to Redox Modulation.J Am Chem Soc. 2025 Feb 26;147(8):6373-6377. doi: 10.1021/jacs.5c00157. Epub 2025 Feb 17. J Am Chem Soc. 2025. PMID: 39957581
-
Chemical Versatility in Catalysis and Inhibition of the Class IIb Histone Deacetylases.Acc Chem Res. 2024 Apr 16;57(8):1135-1148. doi: 10.1021/acs.accounts.3c00801. Epub 2024 Mar 26. Acc Chem Res. 2024. PMID: 38530703 Free PMC article. Review.
-
The Brucella Effector Protein BspF Crotonylates TRIM38 to Inhibit NF-κB and MAPK Signaling Pathway.Int J Mol Sci. 2025 Apr 10;26(8):3573. doi: 10.3390/ijms26083573. Int J Mol Sci. 2025. PMID: 40332097 Free PMC article.
-
Global crotonylatome and GWAS revealed a TaSRT1-TaPGK model regulating wheat cold tolerance through mediating pyruvate.Sci Adv. 2023 May 10;9(19):eadg1012. doi: 10.1126/sciadv.adg1012. Epub 2023 May 10. Sci Adv. 2023. PMID: 37163591 Free PMC article.
-
Regulatory Mechanism of Protein Crotonylation and Its Relationship with Cancer.Cells. 2024 Nov 2;13(21):1812. doi: 10.3390/cells13211812. Cells. 2024. PMID: 39513918 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
