

Synaptic branch stability is mediated by non-enzymatic functions of MEC-17/αTAT1 and ATAT-2
- PMID: 35977998
- PMCID: PMC9385713
- DOI: 10.1038/s41598-022-18333-2
Synaptic branch stability is mediated by non-enzymatic functions of MEC-17/αTAT1 and ATAT-2
Abstract
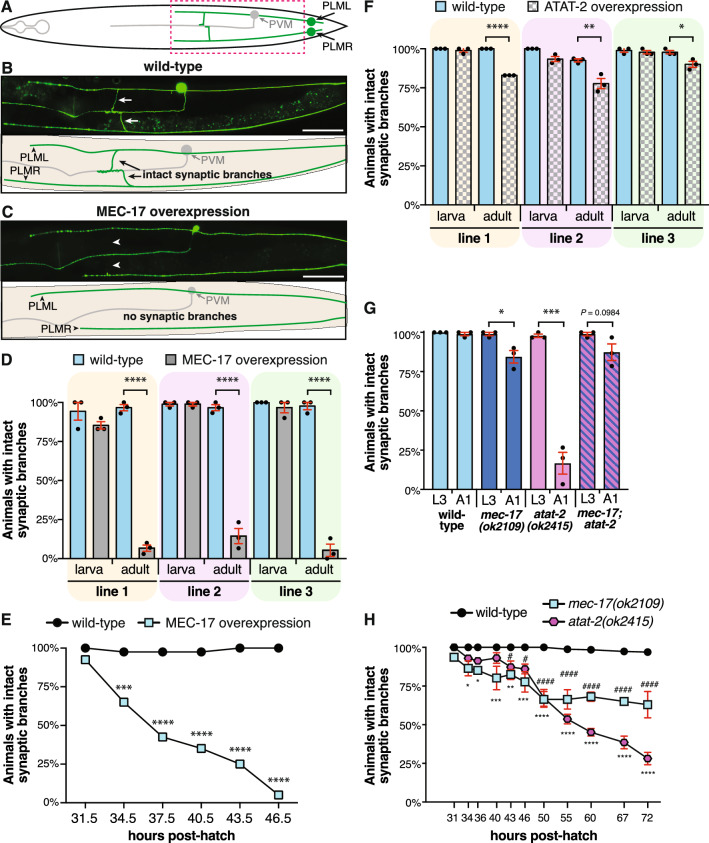
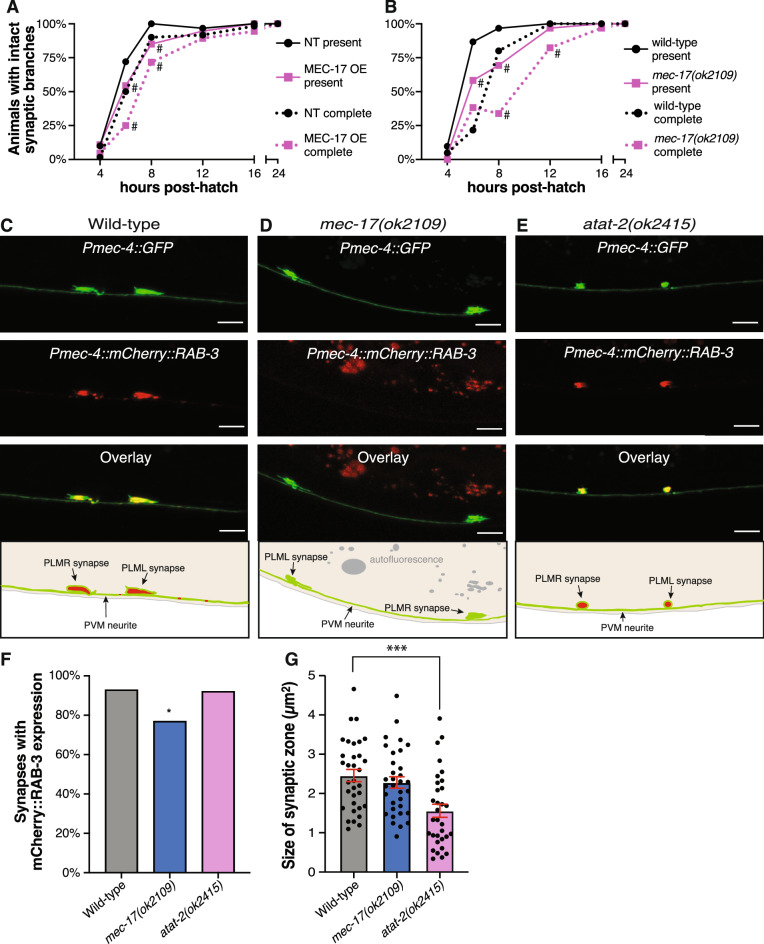
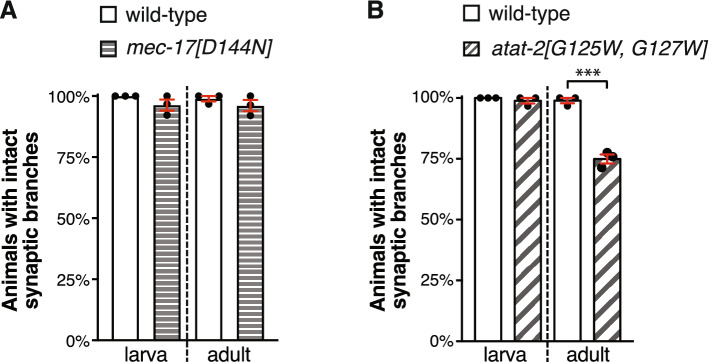
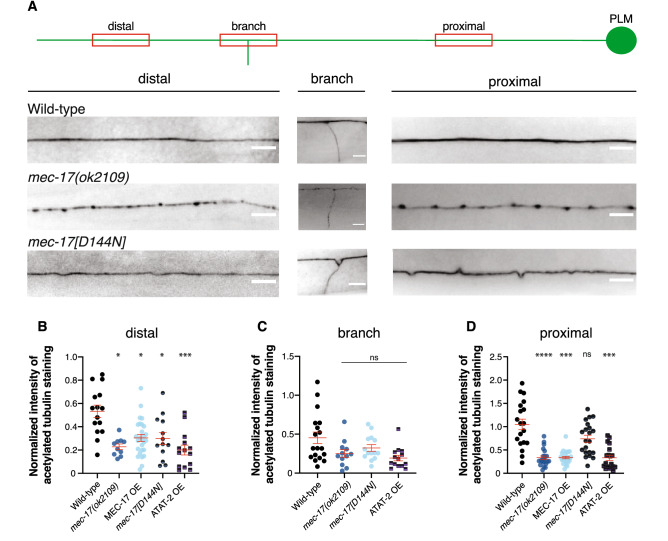
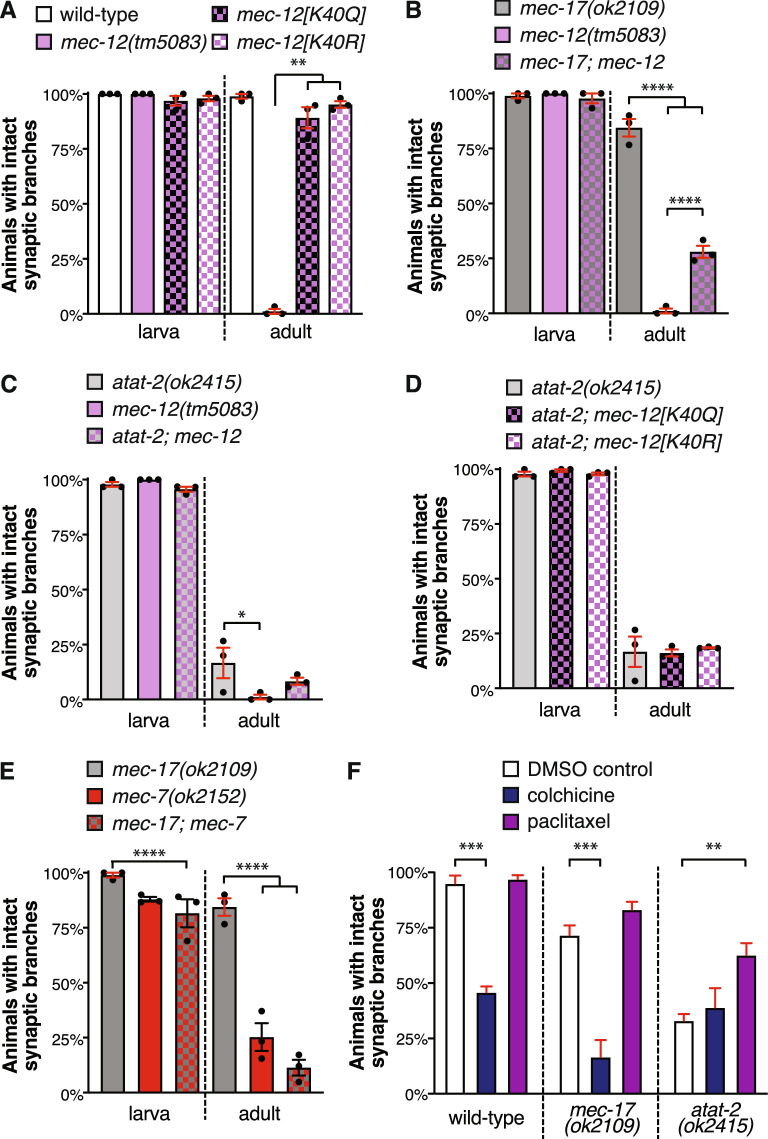
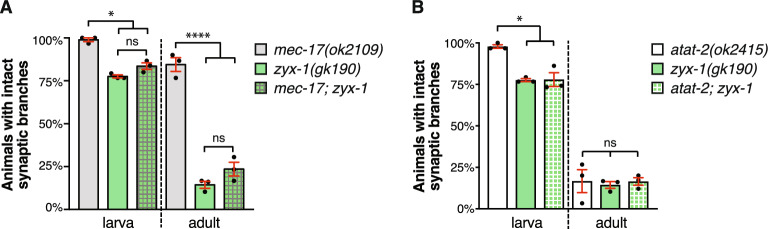
Microtubules are fundamental elements of neuronal structure and function. They are dynamic structures formed from protofilament chains of α- and β-tubulin heterodimers. Acetylation of the lysine 40 (K40) residue of α-tubulin protects microtubules from mechanical stresses by imparting structural elasticity. The enzyme responsible for this acetylation event is MEC-17/αTAT1. Despite its functional importance, however, the consequences of altered MEC-17/αTAT1 levels on neuronal structure and function are incompletely defined. Here we demonstrate that overexpression or loss of MEC-17, or of its functional paralogue ATAT-2, causes a delay in synaptic branch extension, and defective synaptogenesis in the mechanosensory neurons of Caenorhabditis elegans. Strikingly, by adulthood, the synaptic branches in these animals are lost, while the main axon shaft remains mostly intact. We show that MEC-17 and ATAT-2 regulate the stability of the synaptic branches largely independently from their acetyltransferase domains. Genetic analyses reveals novel interactions between both mec-17 and atat-2 with the focal adhesion gene zyx-1/Zyxin, which has previously been implicated in actin remodelling. Together, our results reveal new, acetylation-independent roles for MEC-17 and ATAT-2 in the development and maintenance of neuronal architecture.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Cartoni R, Bradke F, He Z. Enhancing the regeneration of neurons in the central nervous system. Oxf. Res. Encycl. Neurosci. 2019 doi: 10.1093/acrefore/9780190264086.013.217. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
