

GPNMB confers risk for Parkinson's disease through interaction with α-synuclein
- PMID: 35981040
- PMCID: PMC9870036
- DOI: 10.1126/science.abk0637
GPNMB confers risk for Parkinson's disease through interaction with α-synuclein
Abstract
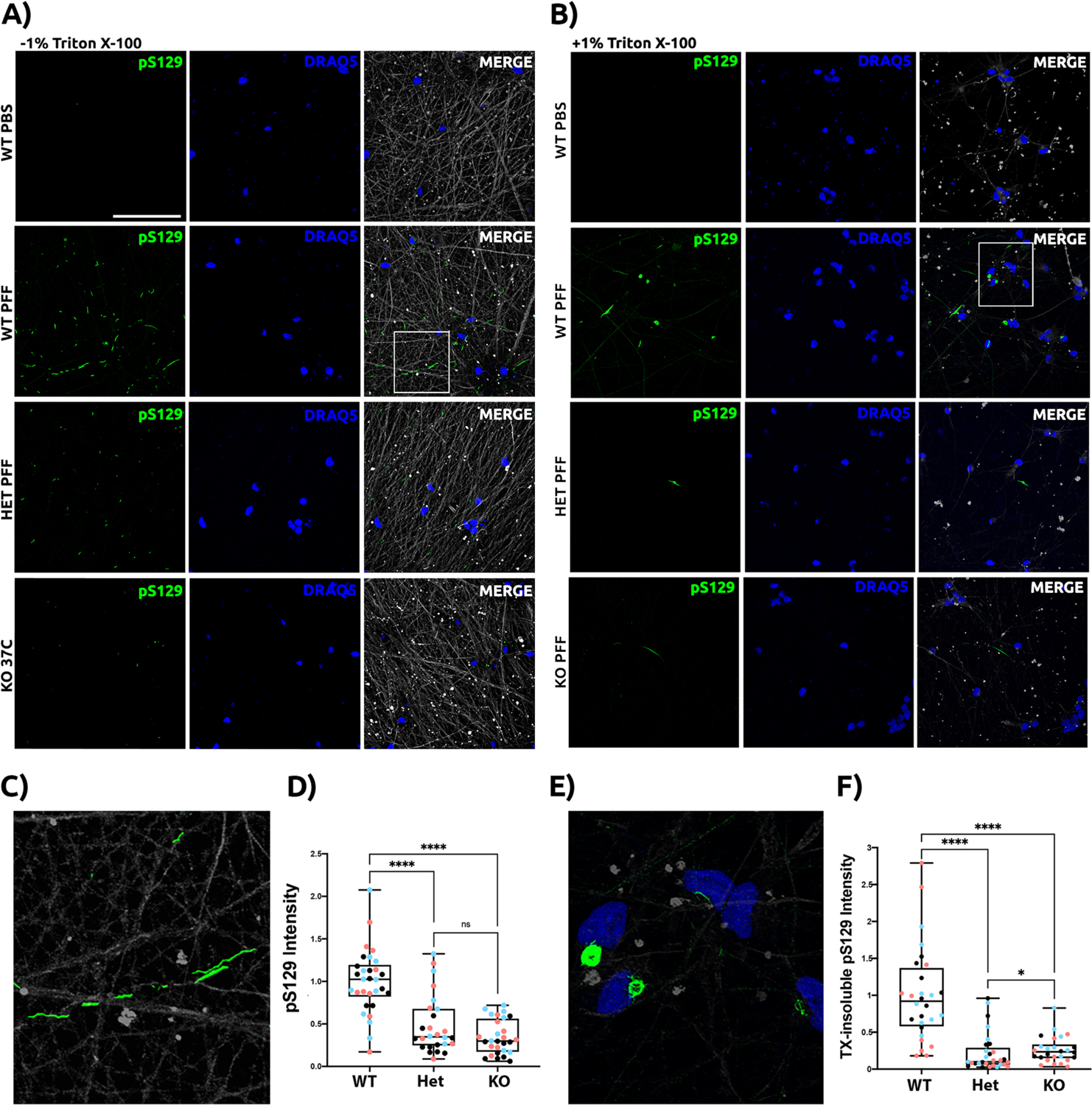
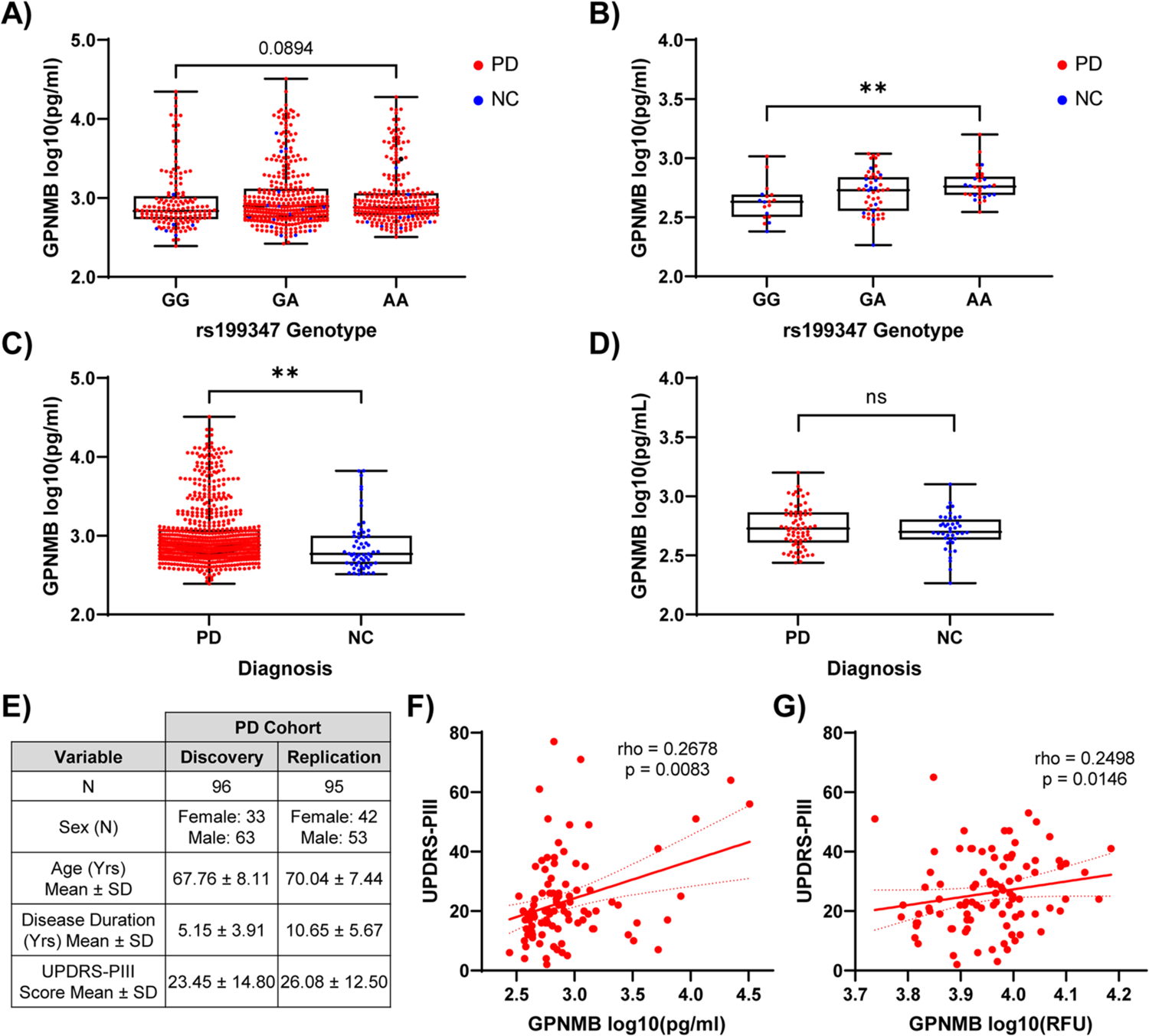
Many risk loci for Parkinson's disease (PD) have been identified by genome-wide association studies (GWASs), but target genes and mechanisms remain largely unknown. We linked the GWAS-derived chromosome 7 locus (sentinel single-nucleotide polymorphism rs199347) to GPNMB through colocalization analyses of expression quantitative trait locus and PD risk signals, confirmed by allele-specific expression studies in the human brain. In cells, glycoprotein nonmetastatic melanoma protein B (GPNMB) coimmunoprecipitated and colocalized with α-synuclein (aSyn). In induced pluripotent stem cell-derived neurons, loss of GPNMB resulted in loss of ability to internalize aSyn fibrils and develop aSyn pathology. In 731 PD and 59 control biosamples, GPNMB was elevated in PD plasma, associating with disease severity. Thus, GPNMB represents a PD risk gene with potential for biomarker development and therapeutic targeting.
Conflict of interest statement
Competing interests:
MDO, YS, and ACP are the inventors of a provisional patent submitted by the University of Pennsylvania that relates to targeting GPNMB as a potential therapeutic in Parkinson’s disease.
Figures
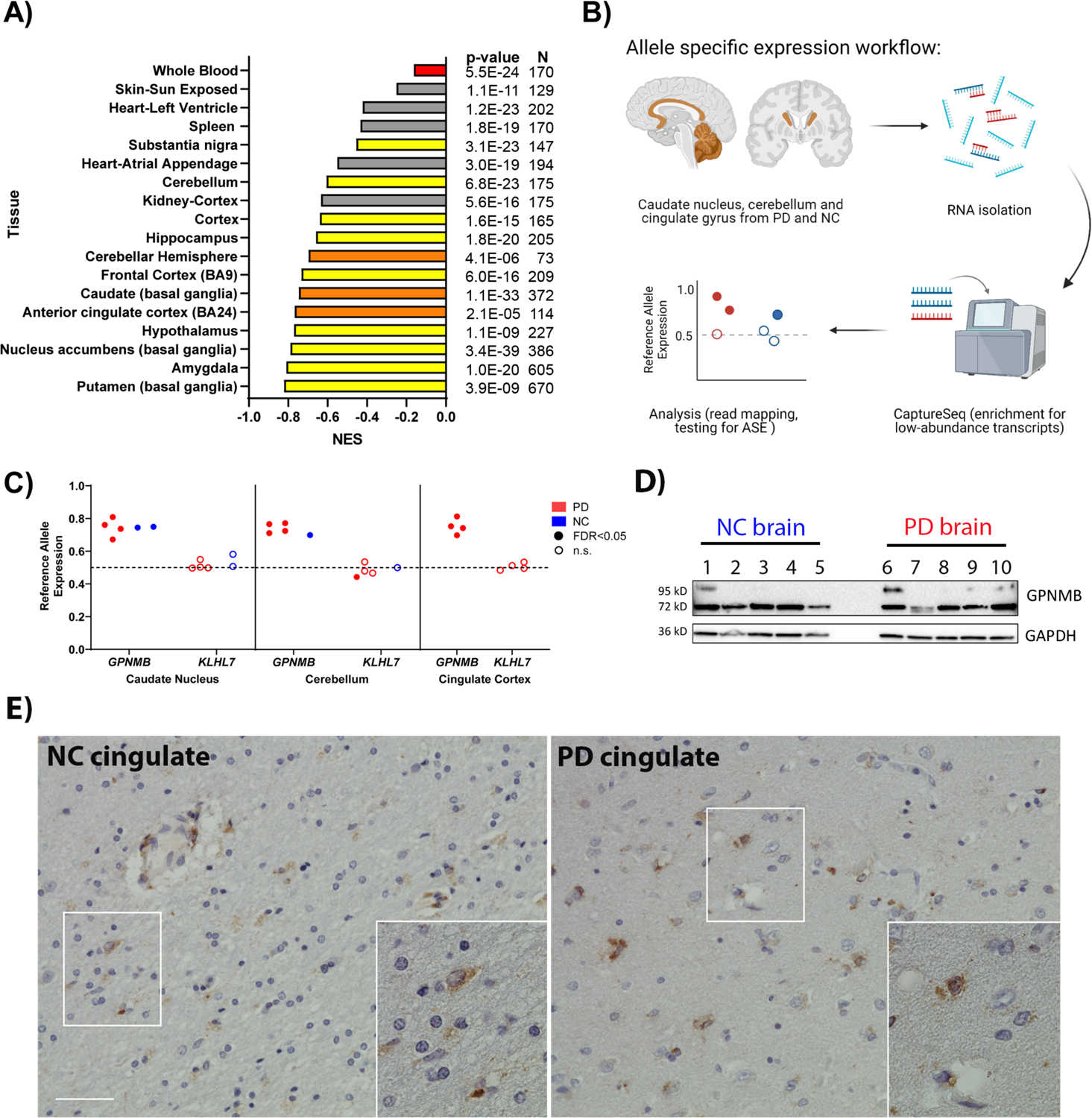
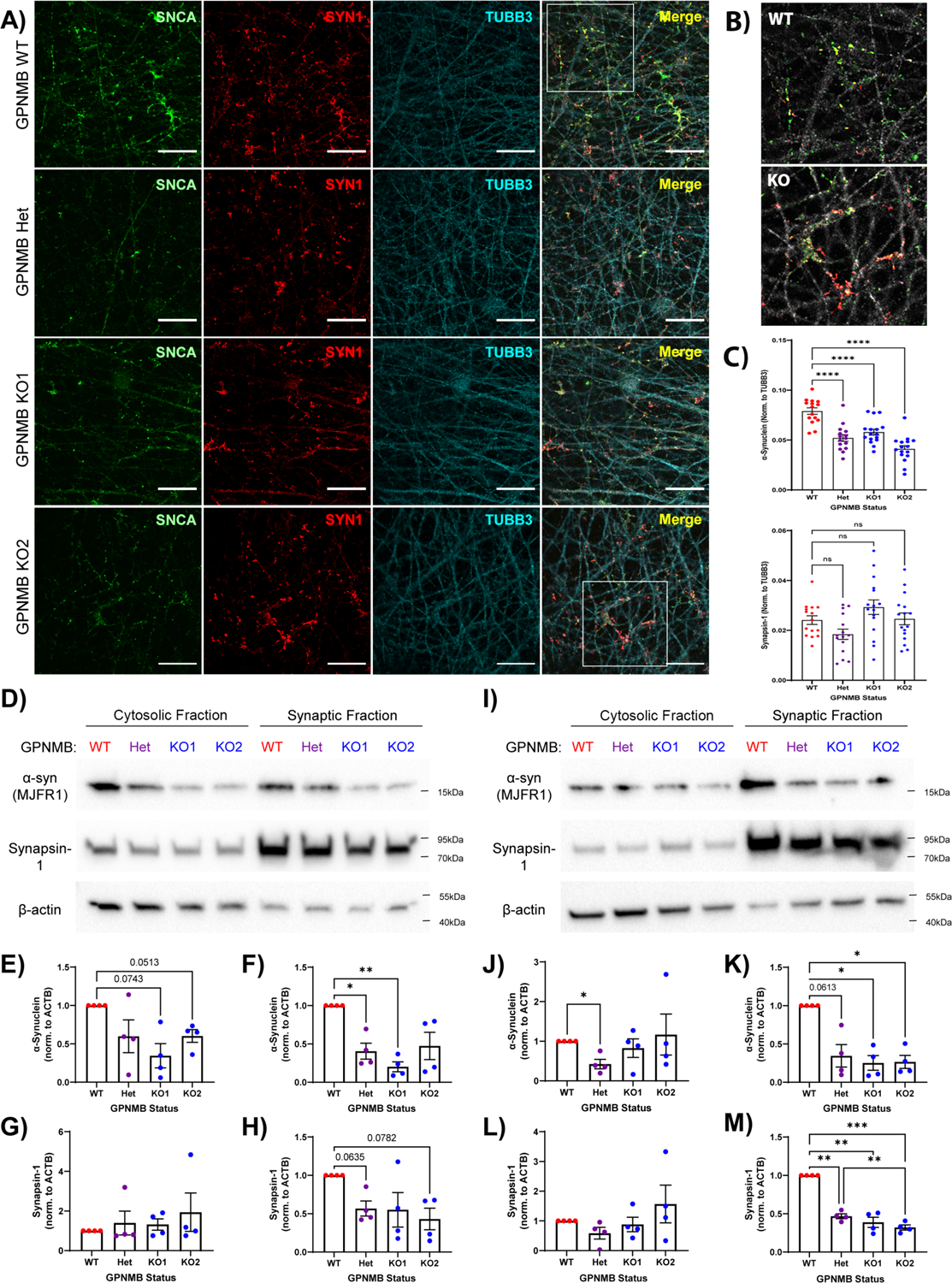
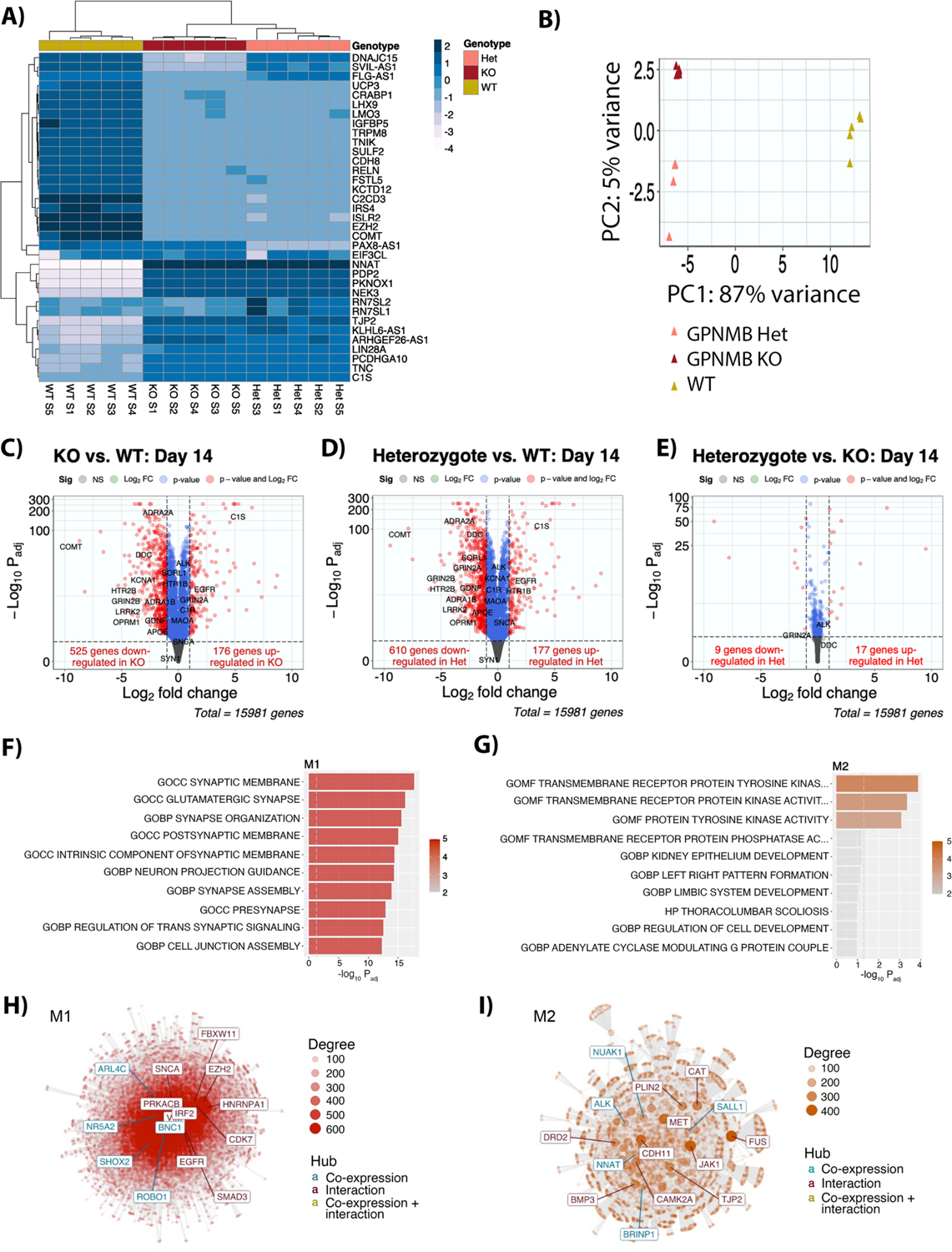
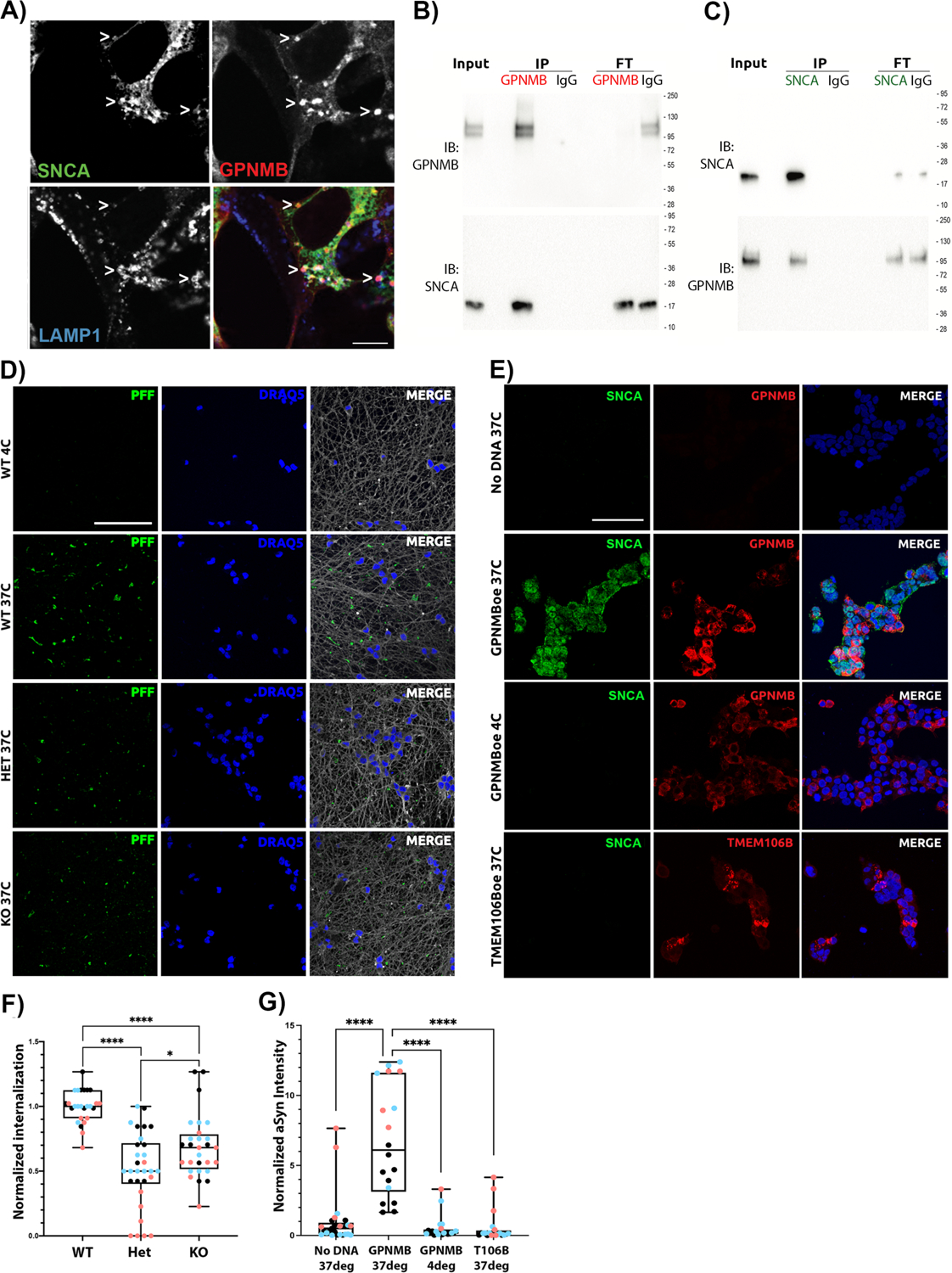
Comment in
-
Toward preventing Parkinson's disease.Science. 2022 Aug 19;377(6608):818-819. doi: 10.1126/science.add7162. Epub 2022 Aug 18. Science. 2022. PMID: 35981039
-
Transmembrane protein interacts with α-synuclein to confer risk of PD.Nat Rev Neurol. 2022 Oct;18(10):576. doi: 10.1038/s41582-022-00716-8. Nat Rev Neurol. 2022. PMID: 36085419 No abstract available.
-
The Glycoprotein GPNMB: A Secret Ingredient for Understanding Parkinson's Disease Etiology?Mov Disord. 2022 Dec;37(12):2342. doi: 10.1002/mds.29264. Epub 2022 Nov 2. Mov Disord. 2022. PMID: 36321268 No abstract available.
-
Linking Genetic Risks to Pathological α-Synuclein Transmission in Parkinson's Disease.Neurosci Bull. 2023 Jul;39(7):1186-1188. doi: 10.1007/s12264-023-01031-y. Epub 2023 Feb 4. Neurosci Bull. 2023. PMID: 36737591 Free PMC article. No abstract available.
References
-
- Dorsey ER, Constantinescu R, Thompson JP, Biglan KM, Holloway RG, Kieburtz K, Marshall FJ, Ravina BM, Schifitto G, Siderowf A, Tanner CM, Projected number of people with Parkinson disease in the most populous nations, 2005 through 2030. Neurology. 68, 384 (2007). - PubMed
-
- Nalls MA, Blauwendraat C, Vallerga CL, Heilbron K, Bandres-Ciga S, Chang D, Tan M, Kia DA, Noyce AJ, Xue A, Bras J, Young E, von Coelln R, Simón-Sánchez J, Schulte C, Sharma M, Krohn L, Pihlstrøm L, Siitonen A, Iwaki H, Leonard H, Faghri F, Gibbs JR, Hernandez DG, Scholz SW, Botia JA, Martinez M, Corvol J-C, Lesage S, Jankovic J, Shulman LM, Sutherland M, Tienari P, Majamaa K, Toft M, Andreassen OA, Bangale T, Brice A, Yang J, Gan-Or Z, Gasser T, Heutink P, Shulman JM, Wood NW, Hinds DA, Hardy JA, Morris HR, Gratten P. M. Visscher, Graham RR, Singleton AB, 23andMe Research Team, System Genomics of Parkinson’s Disease Consortium, International Parkinson’s Disease Genomics Consortium, Identification of novel risk loci, causal insights, and heritable risk for Parkinson’s disease: a meta-analysis of genome-wide association studies. The Lancet Neurology. 18, 1091–1102 (2019). - PMC - PubMed
-
- McFarthing K, Simuni T, Clinical Trial Highlights: Targeting Alpha-Synuclein. Journal of Parkinson’s Disease. 9, 5–16 (2019). - PubMed
-
- Chang D, Nalls MA, Hallgrímsdóttir IB, Hunkapiller J, van der Brug M, Cai F, Kerchner GA, Ayalon G, Bingol B, Sheng M, Hinds D, Behrens TW, Singleton AB, Bhangale TR, Graham RR, I. P. D. G. Consortium, 23andMe Research Team, A meta-analysis of genome-wide association studies identifies 17 new Parkinson’s disease risk loci. Nature Genetics. 49, 1511–1516 (2017). - PMC - PubMed
Supplementary References for Materials and Methods:
-
- Toledo JB, van Deerlin VM, Lee EB, Suh E, Baek Y, Robinson JL, Xie SX, McBride J, Wood EM, Schuck T, Irwin DJ, Gross RG, Hurtig H, McCluskey L, Elman L, Karlawish J, Schellenberg G, Chen-Plotkin A, Wolk D, Grossman M, Arnold SE, Shaw LM, Lee VM-Y, Trojanowski JQ, A platform for discovery: The University of Pennsylvania Integrated Neurodegenerative Disease Biobank. Alzheimer’s & Dementia. 10, 477–484.e1 (2014). - PMC - PubMed
-
- Andrews S, FastQC: A Quality Control Tool for High Throughput Sequence Data [online] (2010), (available at http://www.bioinformatics.babraham.ac.uk/projects/fastqc/).
-
- Dobin A, Gingeras TR, in Data Mining Techniques for the Life Sciences, Carugo O, Eisenhaber F, Eds. (Springer New York, New York, NY, 2016; 10.1007/978-1-4939-3572-7_13), pp. 245–262. - DOI
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
