

A thermometer circuit for hot temperature adjusts Drosophila behavior to persistent heat
- PMID: 35981537
- PMCID: PMC9529852
- DOI: 10.1016/j.cub.2022.07.060
A thermometer circuit for hot temperature adjusts Drosophila behavior to persistent heat
Abstract
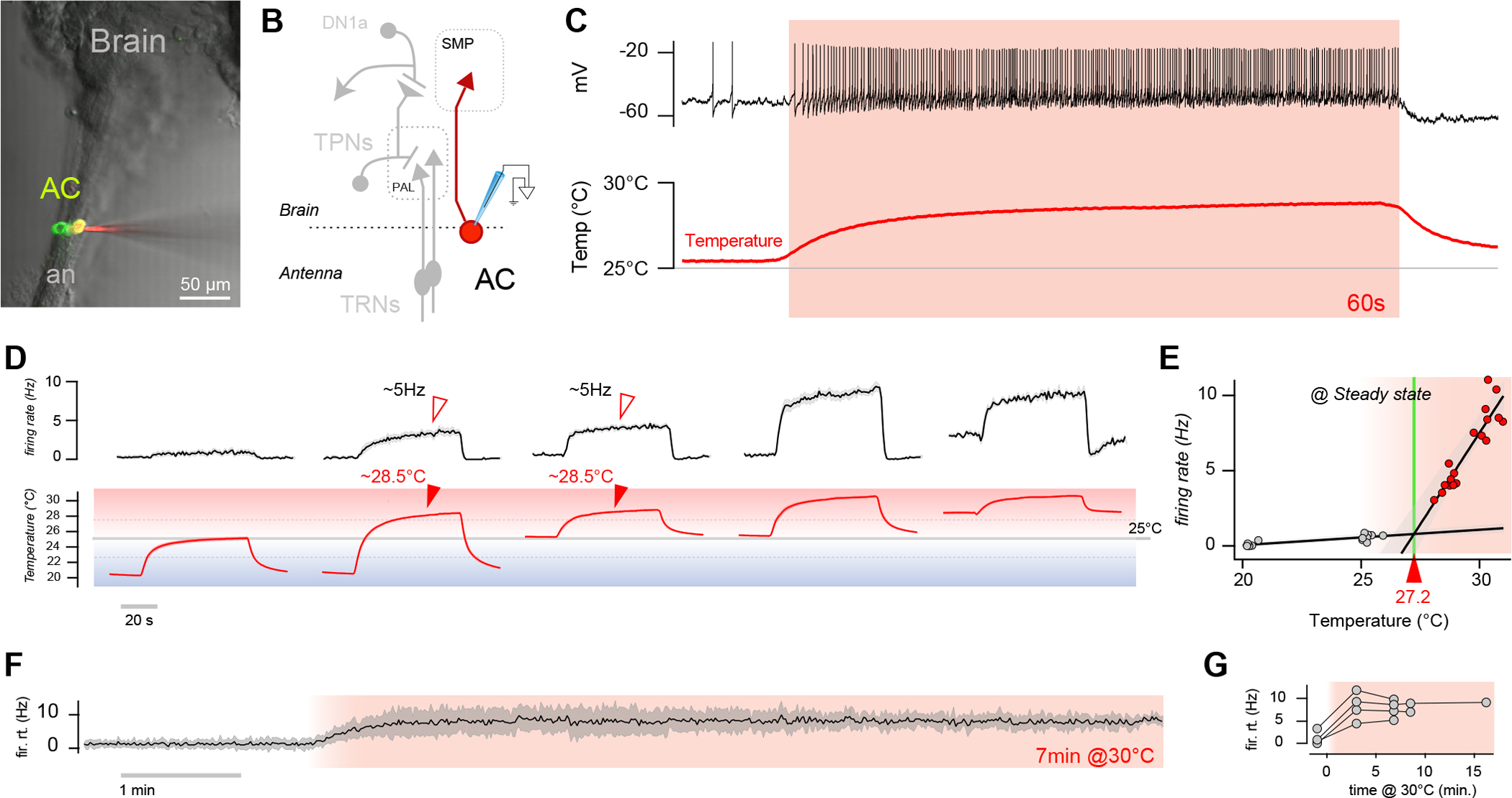
Small poikilotherms such as the fruit fly Drosophila depend on absolute temperature measurements to identify external conditions that are above (hot) or below (cold) their preferred range and to react accordingly. Hot and cold temperatures have a different impact on fly activity and sleep, but the circuits and mechanisms that adjust behavior to specific thermal conditions are not well understood. Here, we use patch-clamp electrophysiology to show that internal thermosensory neurons located within the fly head capsule (the AC neurons1) function as a thermometer active in the hot range. ACs exhibit sustained firing rates that scale with absolute temperature-but only for temperatures above the fly's preferred ∼25°C (i.e., "hot" temperature). We identify ACs in the fly brain connectome and demonstrate that they target a single class of circadian neurons, the LPNs.2 LPNs receive excitatory drive from ACs and respond robustly to hot stimuli, but their responses do not exclusively rely on ACs. Instead, LPNs receive independent drive from thermosensory neurons of the fly antenna via a new class of second-order projection neurons (TPN-IV). Finally, we show that silencing LPNs blocks the restructuring of daytime "siesta" sleep, which normally occurs in response to persistent heat. Our previous work described a distinct thermometer circuit for cold temperature.3 Together, the results demonstrate that the fly nervous system separately encodes and relays absolute hot and cold temperature information, show how patterns of sleep and activity can be adapted to specific temperature conditions, and illustrate how persistent drive from sensory pathways can impact behavior on extended temporal scales.
Keywords: AC neurons; Drosophila; LPNs; circadian rhythms; clock neurons; daytime sleep; electrophysiology; sleep and activity; temperature; thermosensation.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
