

Archetype tasks link intratumoral heterogeneity to plasticity and cancer hallmarks in small cell lung cancer
- PMID: 35981544
- PMCID: PMC9615940
- DOI: 10.1016/j.cels.2022.07.006
Archetype tasks link intratumoral heterogeneity to plasticity and cancer hallmarks in small cell lung cancer
Abstract
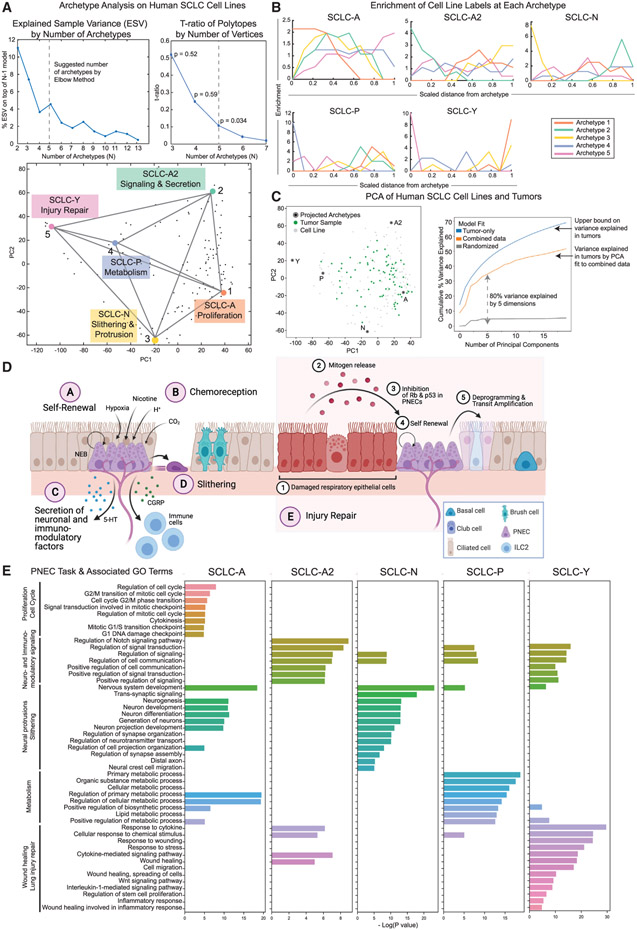
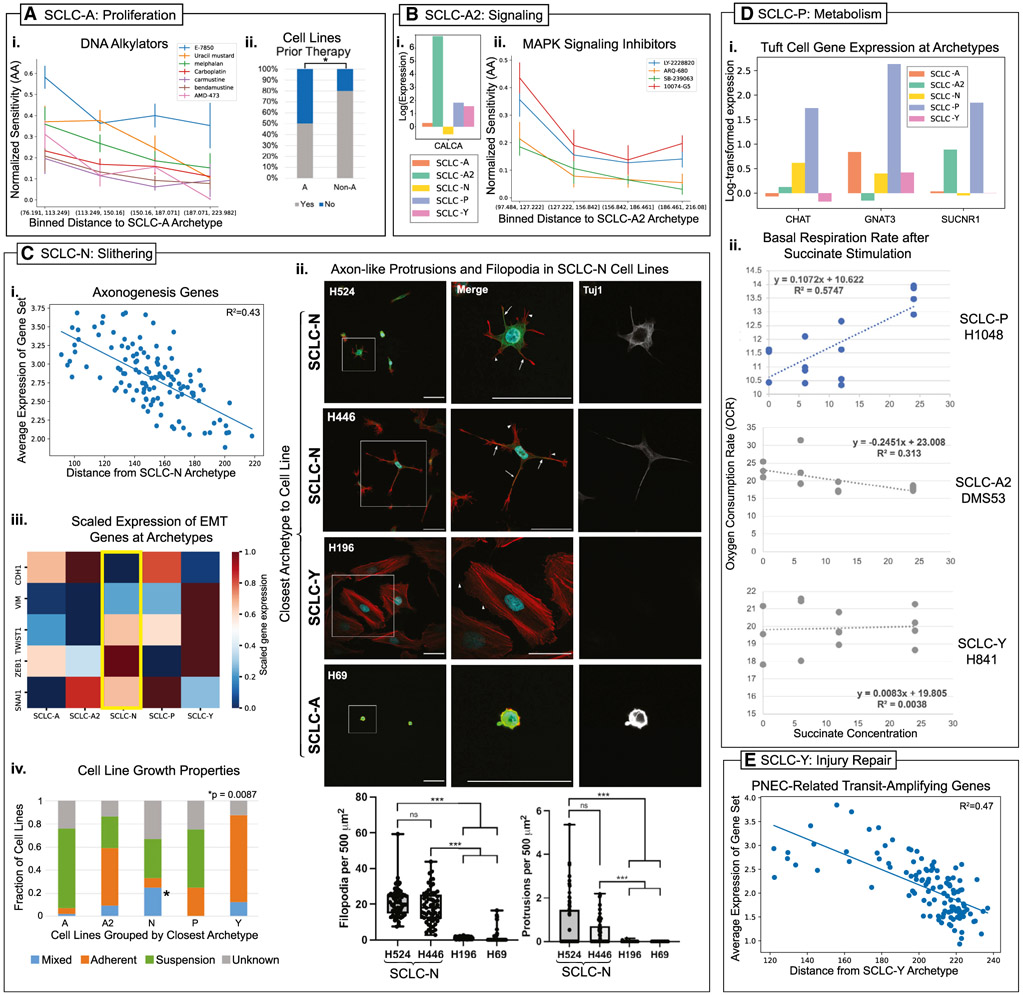
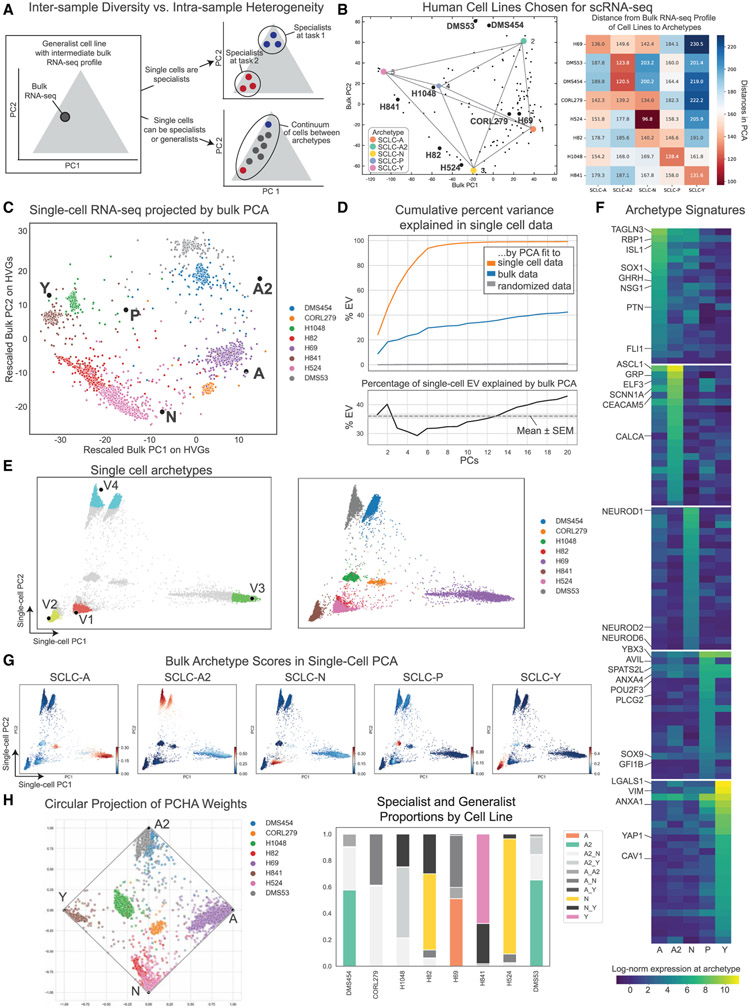
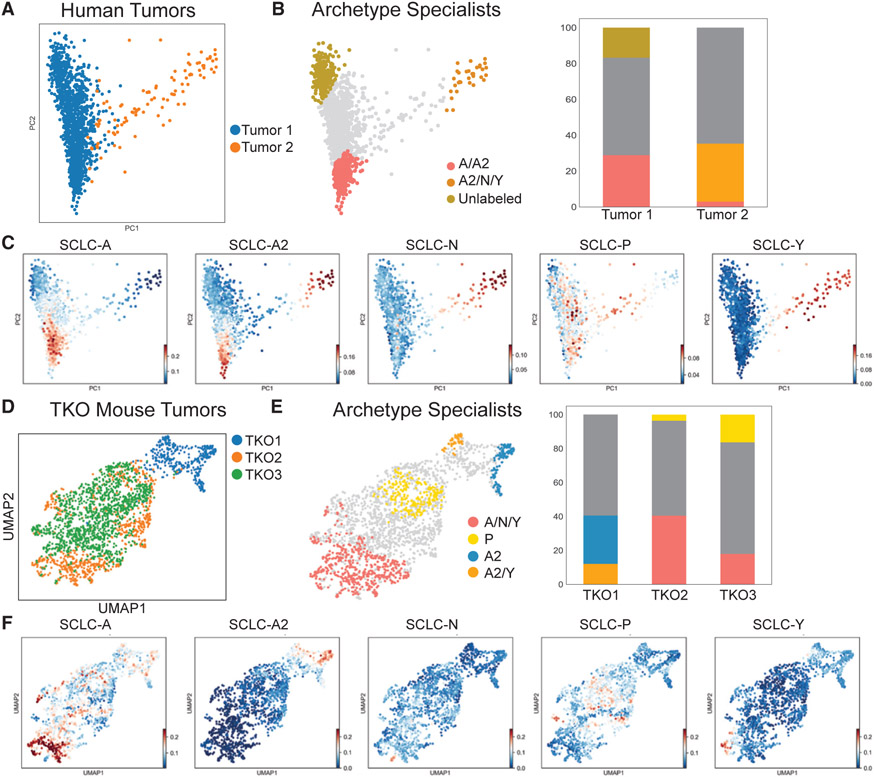
Small cell lung cancer (SCLC) tumors comprise heterogeneous mixtures of cell states, categorized into neuroendocrine (NE) and non-neuroendocrine (non-NE) transcriptional subtypes. NE to non-NE state transitions, fueled by plasticity, likely underlie adaptability to treatment and dismal survival rates. Here, we apply an archetypal analysis to model plasticity by recasting SCLC phenotypic heterogeneity through multi-task evolutionary theory. Cell line and tumor transcriptomics data fit well in a five-dimensional convex polytope whose vertices optimize tasks reminiscent of pulmonary NE cells, the SCLC normal counterparts. These tasks, supported by knowledge and experimental data, include proliferation, slithering, metabolism, secretion, and injury repair, reflecting cancer hallmarks. SCLC subtypes, either at the population or single-cell level, can be positioned in archetypal space by bulk or single-cell transcriptomics, respectively, and characterized as task specialists or multi-task generalists by the distance from archetype vertex signatures. In the archetype space, modeling single-cell plasticity as a Markovian process along an underlying state manifold indicates that task trade-offs, in response to microenvironmental perturbations or treatment, may drive cell plasticity. Stifling phenotypic transitions and plasticity may provide new targets for much-needed translational advances in SCLC. A record of this paper's Transparent Peer Review process is included in the supplemental information.
Keywords: RNA velocity; dynamical systems; gene regulatory networks; heterogeneity; phenotypic plasticity; single cell; small cell lung cancer.
Copyright © 2022. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests C.M.L. is a consultant/advisory board member for Pfizer, Novartis, Astra Zeneca, Genoptix, Sequenom, Ariad, Takeda, Blueprints Medicine, Cepheid, Foundation Medicine, Roche, Achilles Therapeutics, Genentech, Syros, Amgen, EMD Serono, and Eli Lilly and reports receiving commercial research grants from Xcovery, Astra Zeneca, and Novartis. W.T.I. is a consultant/advisory board member for Genentech, Jazz Pharma, G1 Therapeutics, Mirati, OncLive, Clinical Care Options, Chardan, Outcomes Insights, Cello Health, and Curio Science. T.G.O. is a consultant/advisory board member for Known Medicine. J.S. receives research funding from Pfizer. V.Q. is an Academic co-Founder and equity holder for Parthenon Therapeutics, Inc. and Duet BioSystems, Inc.
Figures
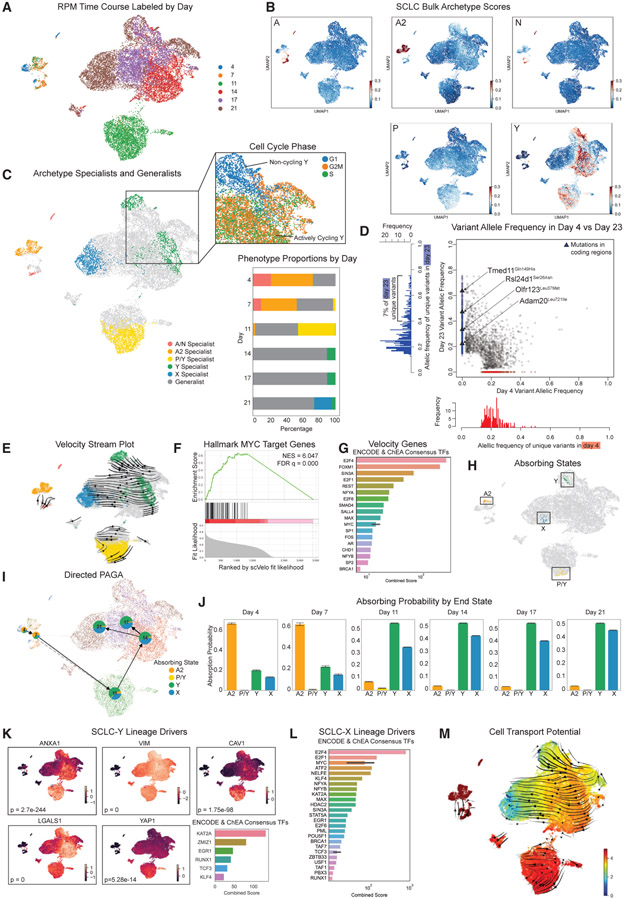
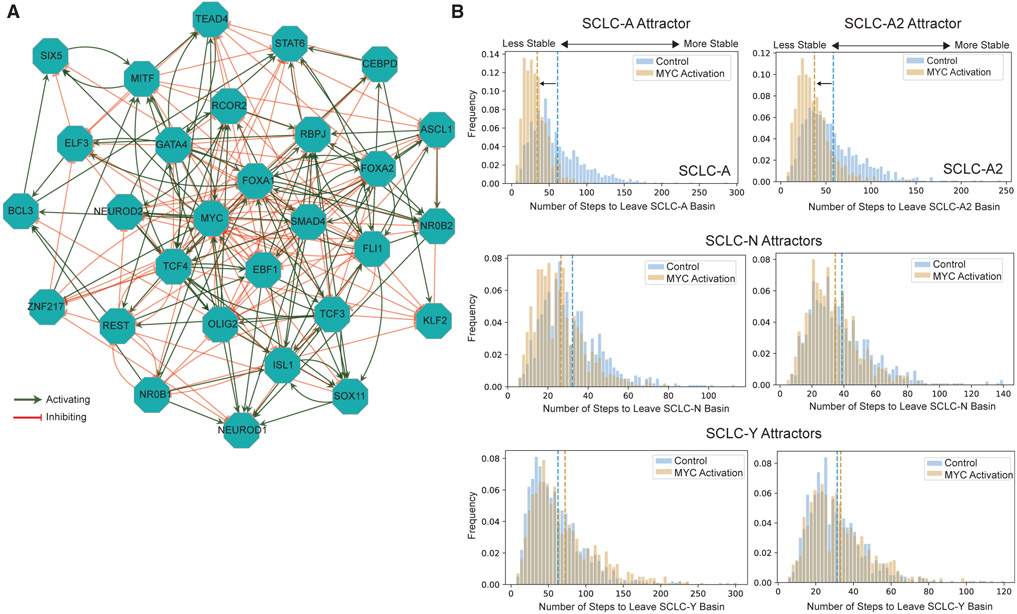
Comment in
-
Unraveling phenotypic plasticity and evolution in small cell lung cancer.Cell Syst. 2022 Sep 21;13(9):687-689. doi: 10.1016/j.cels.2022.08.005. Cell Syst. 2022. PMID: 36137510
References
-
- Agaimy A, Erlenbach-Wünsch K, Konukiewitz B, Schmitt AM, Rieker RJ, Vieth M, Kiesewetter F, Hartmann A, Zamboni G, Perren A, and Klöppel G (2013). ISL1 expression is not restricted to pancreatic well-differentiated neuroendocrine neoplasms, but is also commonly found in well and poorly differentiated neuroendocrine neoplasms of extrapancreatic origin. Mod. Pathol 26, 995–1003. - PubMed
Publication types
MeSH terms
Grants and funding
- U01 CA224276/CA/NCI NIH HHS/United States
- U24 CA213274/CA/NCI NIH HHS/United States
- K12 CA090625/CA/NCI NIH HHS/United States
- R01 DK106228/DK/NIDDK NIH HHS/United States
- U01 CA231844/CA/NCI NIH HHS/United States
- R50 CA243783/CA/NCI NIH HHS/United States
- UG1 CA233259/CA/NCI NIH HHS/United States
- P50 CA236733/CA/NCI NIH HHS/United States
- U01 CA215845/CA/NCI NIH HHS/United States
- R01 CA251147/CA/NCI NIH HHS/United States
- U01 CA215798/CA/NCI NIH HHS/United States
- U54 CA217450/CA/NCI NIH HHS/United States
- K00 CA234920/CA/NCI NIH HHS/United States
- R01 CA217987/CA/NCI NIH HHS/United States
- R01 DK103831/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
