

RNA Pol II pausing facilitates phased pluripotency transitions by buffering transcription
- PMID: 35981753
- PMCID: PMC9480856
- DOI: 10.1101/gad.349565.122
RNA Pol II pausing facilitates phased pluripotency transitions by buffering transcription
Abstract
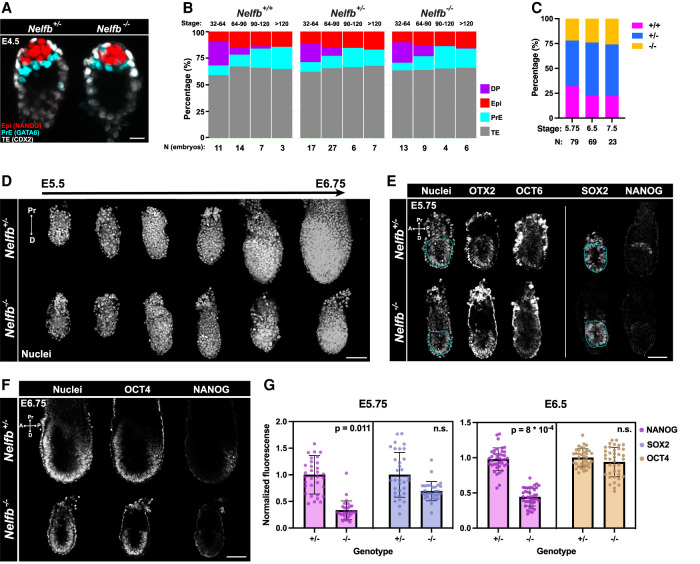
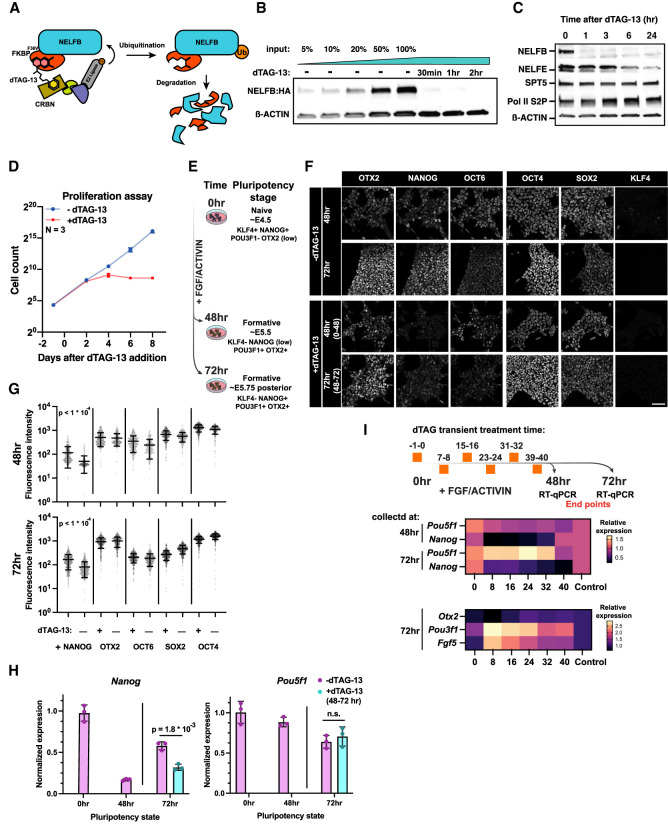
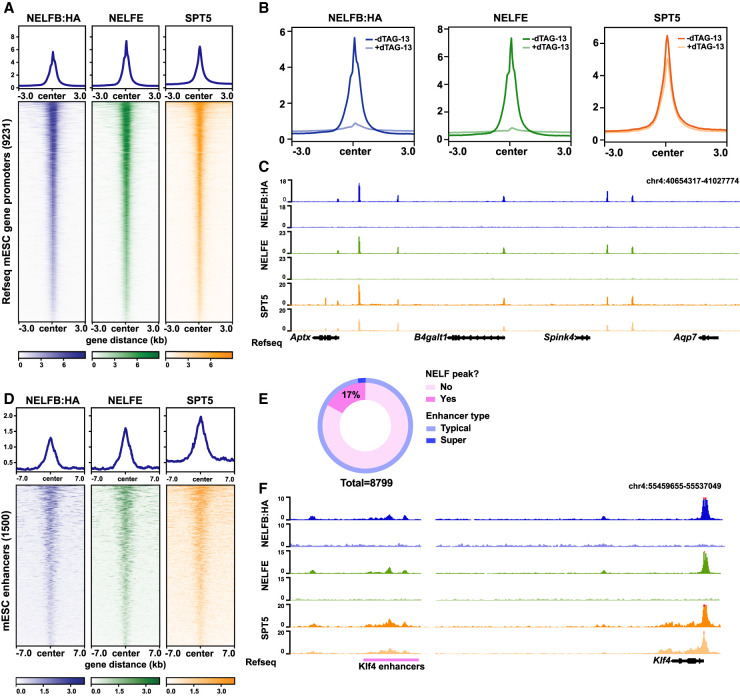
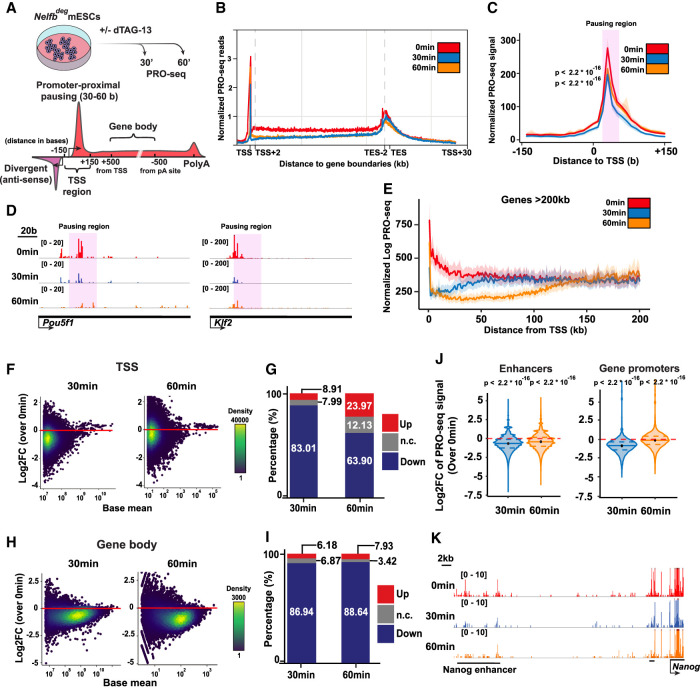
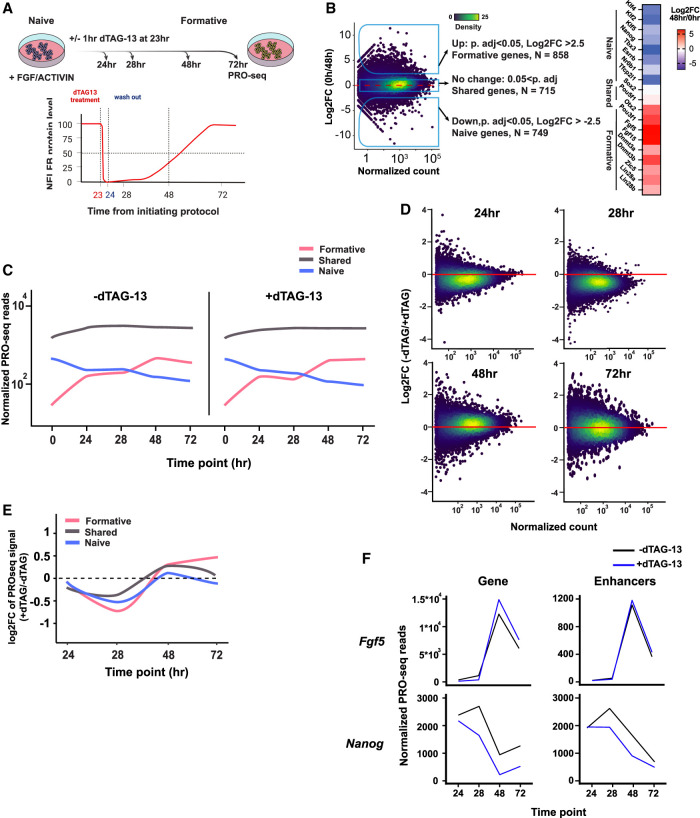
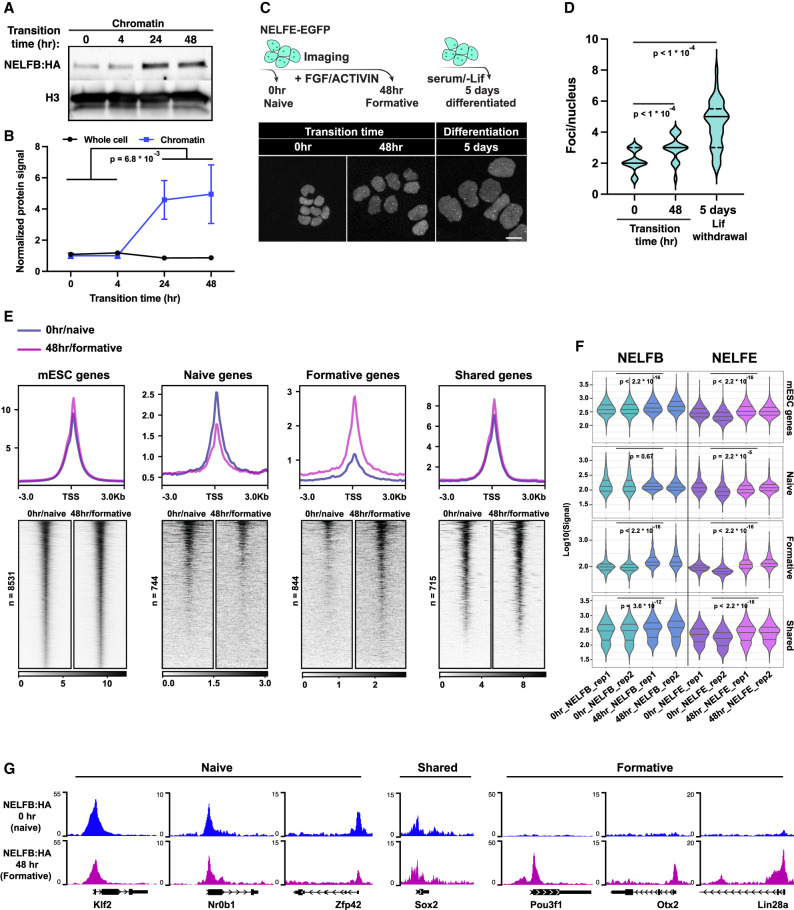
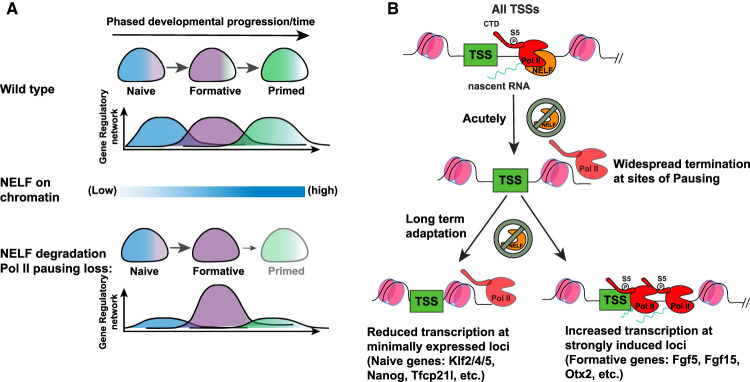
Promoter-proximal RNA Pol II pausing is a critical step in transcriptional control. Pol II pausing has been predominantly studied in tissue culture systems. While Pol II pausing has been shown to be required for mammalian development, the phenotypic and mechanistic details of this requirement are unknown. Here, we found that loss of Pol II pausing stalls pluripotent state transitions within the epiblast of the early mouse embryo. Using Nelfb -/- mice and a NELFB degron mouse pluripotent stem cell model, we show that embryonic stem cells (ESCs) representing the naïve state of pluripotency successfully initiate a transition program but fail to balance levels of induced and repressed genes and enhancers in the absence of NELF. We found an increase in chromatin-associated NELF during transition from the naïve to later pluripotent states. Overall, our work defines the acute and long-term molecular consequences of NELF loss and reveals a role for Pol II pausing in the pluripotency continuum as a modulator of cell state transitions.
Keywords: NELF; dTAG; degron; embryonic stem cells; epiblast; mouse embryo; pausing; pluripotency; transcription.
© 2022 Abuhashem et al.; Published by Cold Spring Harbor Laboratory Press.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials