

STUB1 is an intracellular checkpoint for interferon gamma sensing
- PMID: 35982220
- PMCID: PMC9388626
- DOI: 10.1038/s41598-022-18404-4
STUB1 is an intracellular checkpoint for interferon gamma sensing
Abstract
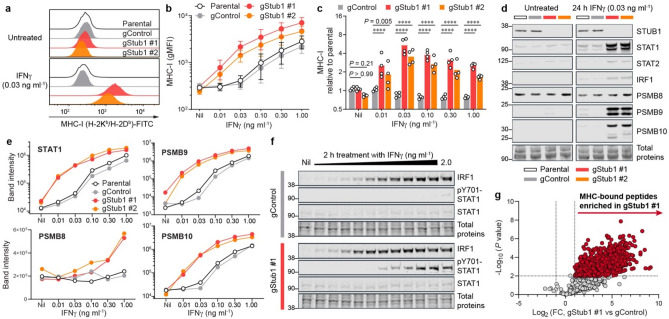
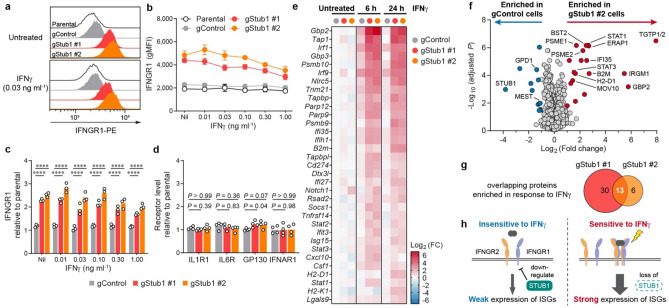
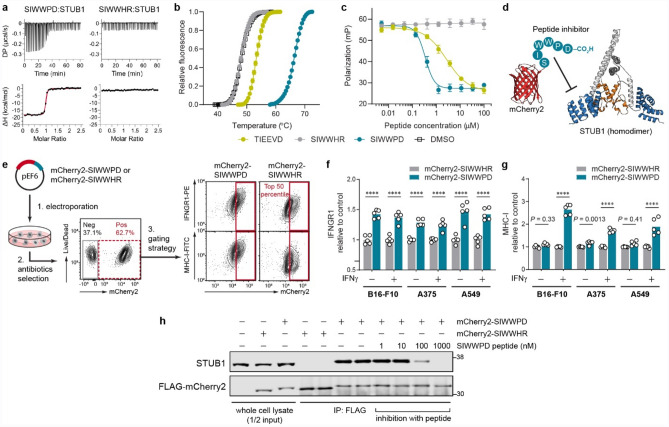
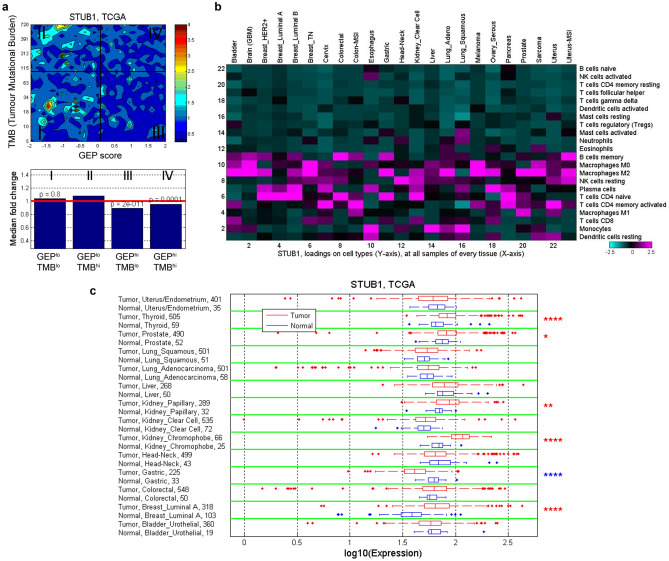
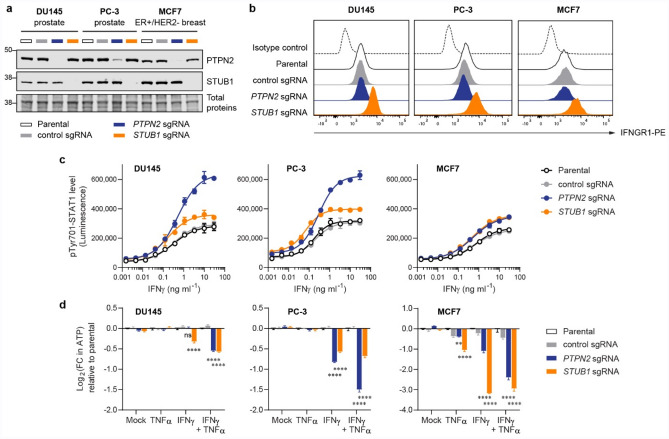
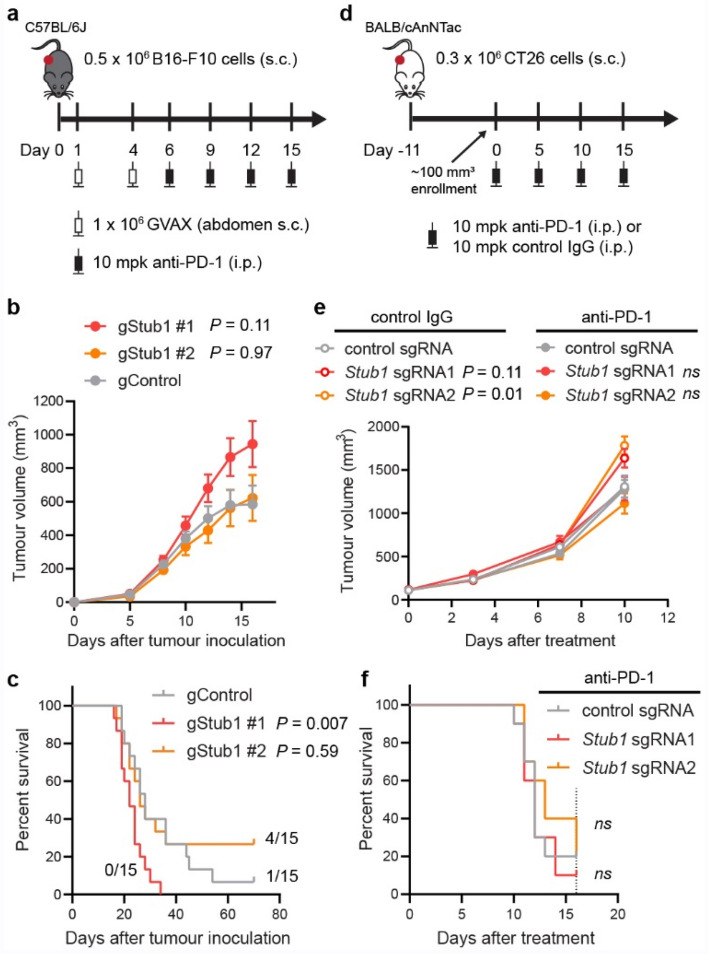
Immune checkpoint blockade (ICB) leads to durable and complete tumour regression in some patients but in others gives temporary, partial or no response. Accordingly, significant efforts are underway to identify tumour-intrinsic mechanisms underlying ICB resistance. Results from a published CRISPR screen in a mouse model suggested that targeting STUB1, an E3 ligase involved in protein homeostasis, may overcome ICB resistance but the molecular basis of this effect remains unclear. Herein, we report an under-appreciated role of STUB1 to dampen the interferon gamma (IFNγ) response. Genetic deletion of STUB1 increased IFNGR1 abundance on the cell surface and thus enhanced the downstream IFNγ response as showed by multiple approaches including Western blotting, flow cytometry, qPCR, phospho-STAT1 assay, immunopeptidomics, proteomics, and gene expression profiling. Human prostate and breast cancer cells with STUB1 deletion were also susceptible to cytokine-induced growth inhibition. Furthermore, blockade of STUB1 protein function recapitulated the STUB1-null phenotypes. Despite these encouraging in vitro data and positive implications from clinical datasets, we did not observe in vivo benefits of inactivating Stub1 in mouse syngeneic tumour models-with or without combination with anti-PD-1 therapy. However, our findings elucidate STUB1 as a barrier to IFNγ sensing, prompting further investigations to assess if broader inactivation of human STUB1 in both tumors and immune cells could overcome ICB resistance.
© 2022. The Author(s).
Conflict of interest statement
All authors are current or former employees of subsidiaries of Merck & Co., Inc., Kenilworth, NJ, USA.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
