

A biosynthetic pathway for the selective sulfonation of steroidal metabolites by human gut bacteria
- PMID: 35982310
- PMCID: PMC10327491
- DOI: 10.1038/s41564-022-01176-y
A biosynthetic pathway for the selective sulfonation of steroidal metabolites by human gut bacteria
Abstract
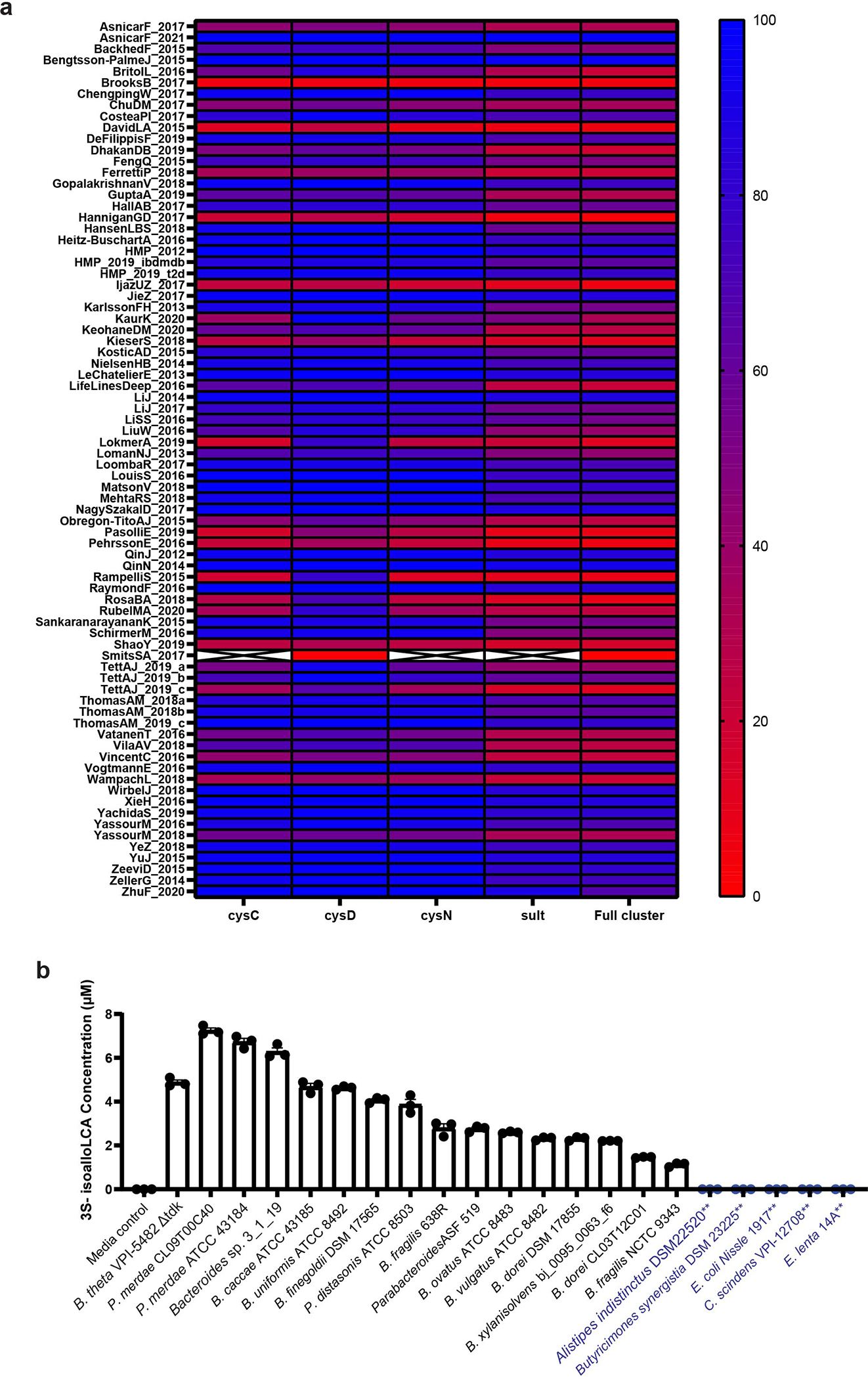
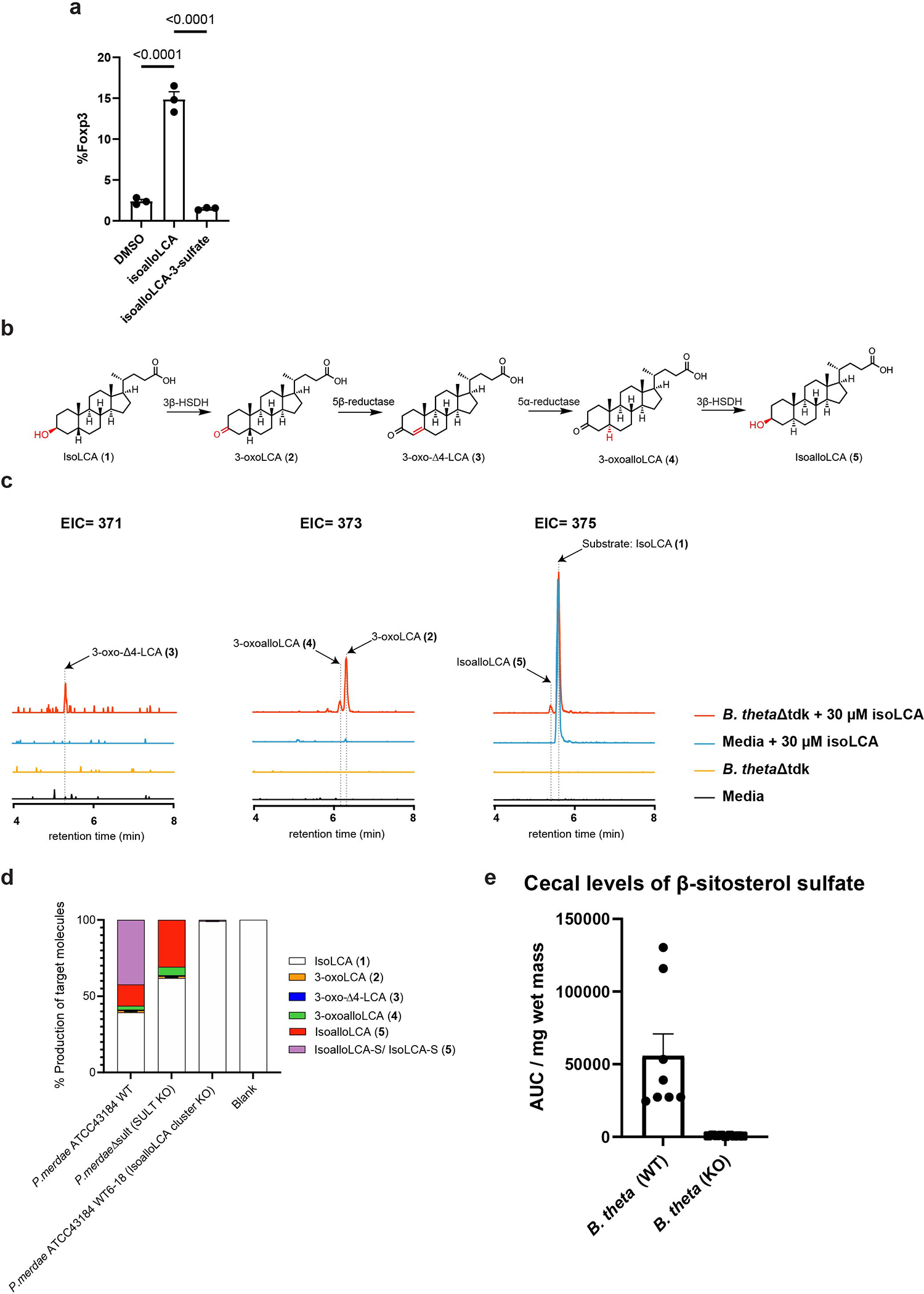
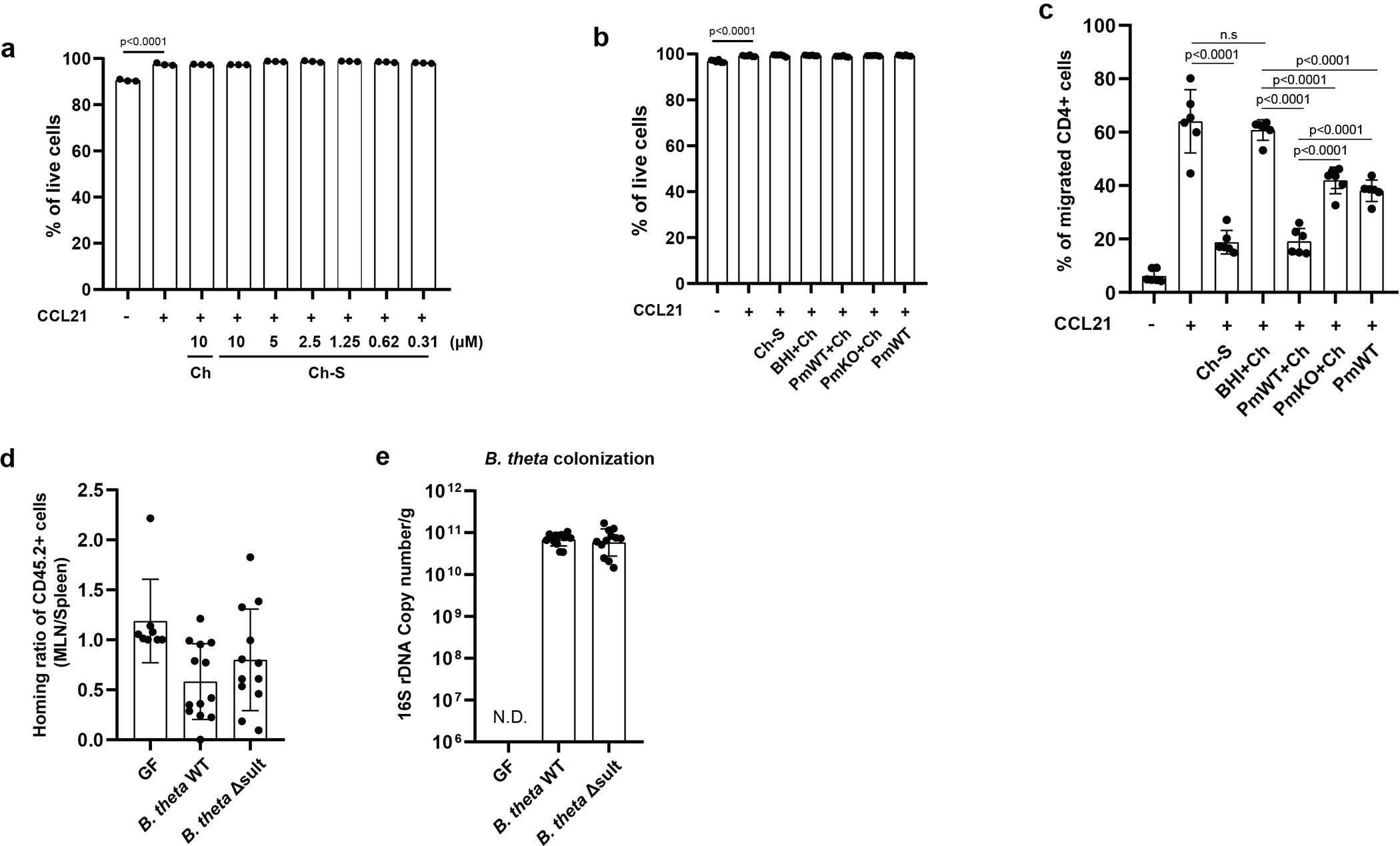
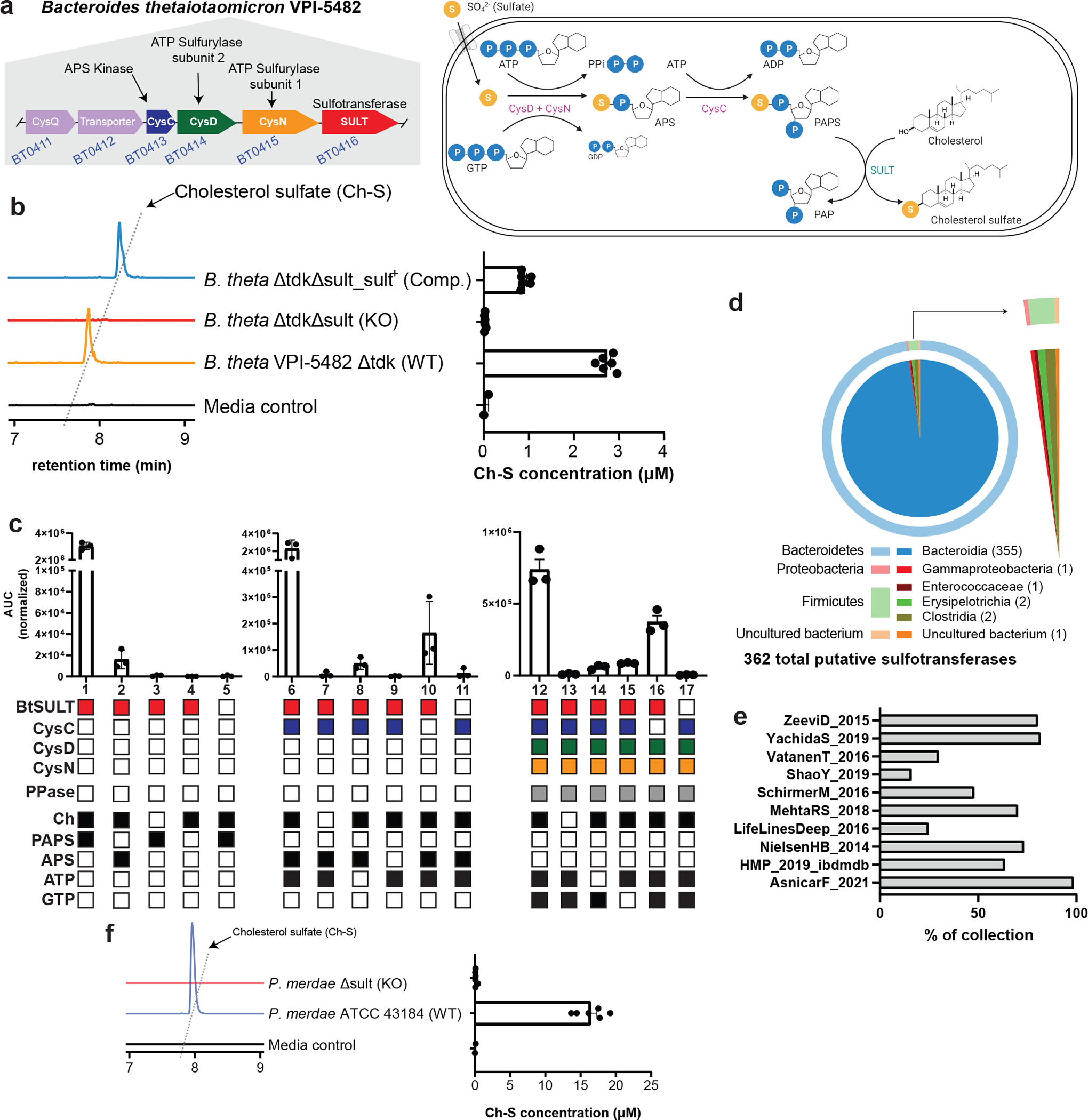
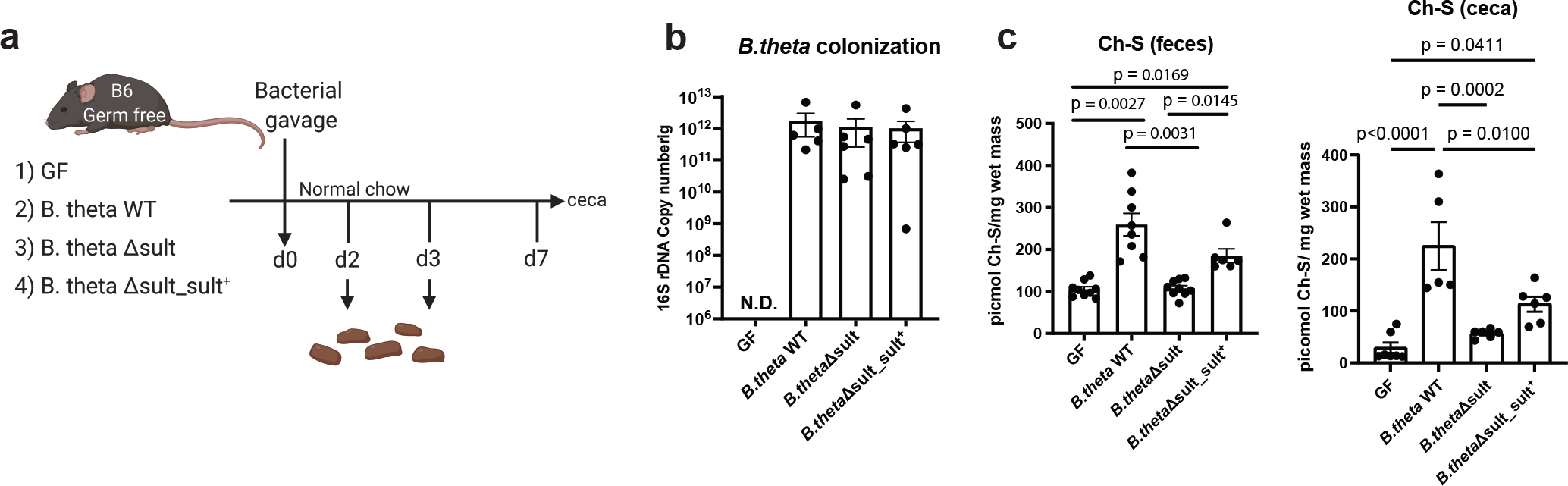
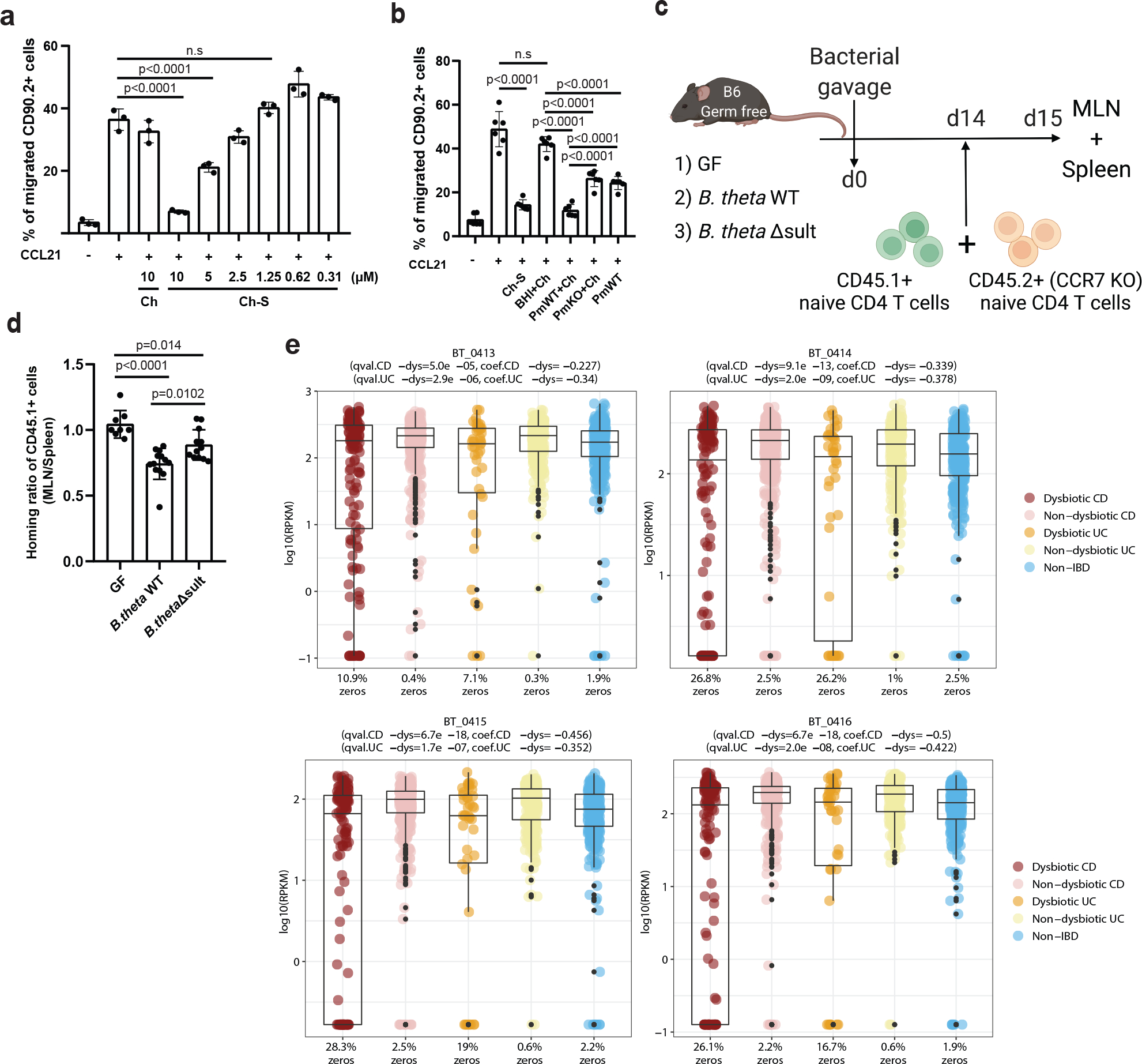
Members of the human gut microbiome enzymatically process many bioactive molecules in the gastrointestinal tract. Most gut bacterial modifications characterized so far are hydrolytic or reductive in nature. Here we report that abundant human gut bacteria from the phylum Bacteroidetes perform conjugative modifications by selectively sulfonating steroidal metabolites. While sulfonation is a ubiquitous biochemical modification, this activity has not yet been characterized in gut microbes. Using genetic and biochemical approaches, we identify a widespread biosynthetic gene cluster that encodes both a sulfotransferase (BtSULT, BT0416) and enzymes that synthesize the sulfonate donor adenosine 3'-phosphate-5'-phosphosulfate (PAPS), including an APS kinase (CysC, BT0413) and an ATP sulfurylase (CysD and CysN, BT0414-BT0415). BtSULT selectively sulfonates steroidal metabolites with a flat A/B ring fusion, including cholesterol. Germ-free mice monocolonized with Bacteroides thetaiotaomicron ΔBT0416 exhibited reduced gastrointestinal levels of cholesterol sulfate (Ch-S) compared with wild-type B. thetaiotaomicron-colonized mice. The presence of BtSULT and BtSULT homologues in bacteria inhibited leucocyte migration in vitro and in vivo, and abundances of cluster genes were significantly reduced in patients with inflammatory bowel disease. Together, these data provide a mechanism by which gut bacteria sulfonate steroidal metabolites and suggest that these compounds can modulate immune cell trafficking in the host.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Competing interests
A.S.D. is a consultant for Takeda Pharmaceuticals and Axial Therapeutics. J.R.H. is a consultant for CJ Research Center, LLC. C.H. is on the scientific advisory boards of Seres Therapeutics, Empress Therapeutics, and ZOE Nutrition. The remaining authors declare no competing interests.
Figures









Comment in
-
Microbial regulation of cholesterol homeostasis.Nat Microbiol. 2022 Sep;7(9):1327-1328. doi: 10.1038/s41564-022-01186-w. Nat Microbiol. 2022. PMID: 35982309 No abstract available.
References
-
- Ervin SM et al. Structural Insights into Endobiotic Reactivation by Human Gut Microbiome-Encoded Sulfatases. Biochemistry-us 59, 3939–3950 (2020). - PubMed
-
- Jancova P, Anzenbacher P & Anzenbacherova E PHASE II DRUG METABOLIZING ENZYMES. Biomed Pap 154, 103–116 (2010). - PubMed
-
- Lindsay J, Wang LL, Li Y & Zhou SF Structure, function and polymorphism of human cytosolic sulfotransferases. Curr Drug Metab 9, 99–105 (2008). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
