

RNAi-mediated knockdown of gut receptor-like genes prohibitin and α-amylase altered the susceptibility of Galleria mellonella to Cry1AcF toxin
- PMID: 35982422
- PMCID: PMC9389788
- DOI: 10.1186/s12864-022-08843-8
RNAi-mediated knockdown of gut receptor-like genes prohibitin and α-amylase altered the susceptibility of Galleria mellonella to Cry1AcF toxin
Abstract
Background: Due to the prolonged usage of Bt-based biopesticides and Bt-transgenic crops worldwide, insects are continually developing resistance against Cry toxins. This resistance may occur if any mechanistic step in the insecticidal process is disrupted possibly because of the alteration in Cry-receptor binding affinity due to mutation in receptor genes. Compared to other lepidopteran insects, Cry receptor-related research has made asymmetric progress in the model insect Galleria mellonella.
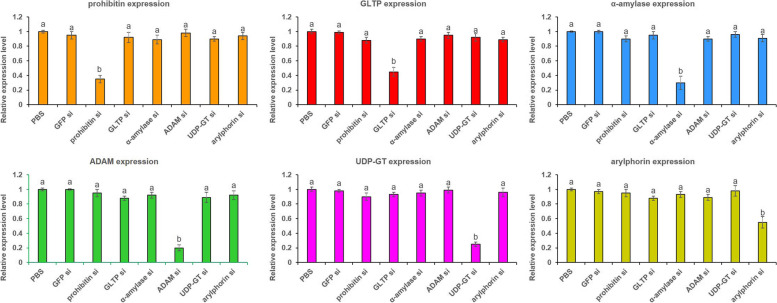
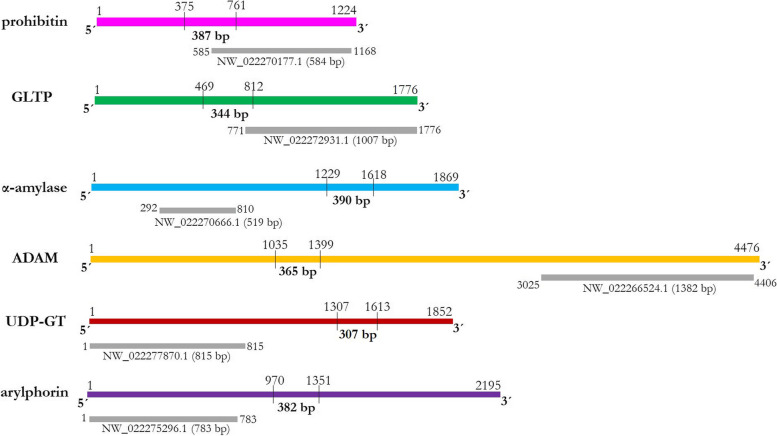
Results: Present study describes the molecular characterization and functional analysis of five Cry toxin receptor-related genes (prohibitin, GLTP, α-amylase, ADAM and UDP-GT) and a gut repair gene (arylphorin) from the gut tissues of G. mellonella. Protein-protein docking analysis revealed that Cry1AcF putatively binds with all the five candidate proteins, suggesting their receptor-like function. These receptor-like genes were significantly overexpressed in the gut tissues of fourth-instar G. mellonella larvae upon early exposure to a sub-lethal dose of Cry1AcF toxin. However, targeted knockdown (by using bacterially-expressed dsRNAs) of these genes led to variable effect on insect susceptibility to Cry1AcF toxin. Insects pre-treated with prohibitin and α-amylase dsRNA exhibited significant reduction in Cry1AcF-induced mortality, suggesting their probable role as Cry receptor. By contrast, insects pre-treated with GLTP, ADAM and UDP-GT dsRNA exhibited no significant decline in mortality. This maybe explained by the possibility of RNAi feedback regulation (as few of the receptors belong to multigene family) or redundant role of GLTP, ADAM and UDP-GT in Cry intoxication process.
Conclusion: Since the laboratory culture of G. mellonella develop Bt resistance quite rapidly, findings of the current investigation may provide some useful information for future Cry receptor-related research in the model insect.
Keywords: ADAM; Cry1AcF; GLTP; Prohibitin; RNAi; UDP-GT; α-amylase.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures






References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
