

Sexual Signals Persist over Deep Time: Ancient Co-option of Bioluminescence for Courtship Displays in Cypridinid Ostracods
- PMID: 35984328
- PMCID: PMC10448971
- DOI: 10.1093/sysbio/syac057
Sexual Signals Persist over Deep Time: Ancient Co-option of Bioluminescence for Courtship Displays in Cypridinid Ostracods
Abstract
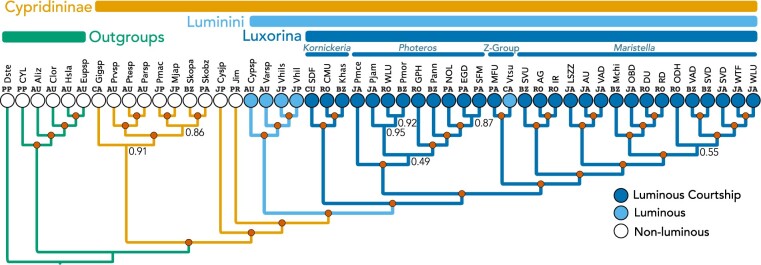
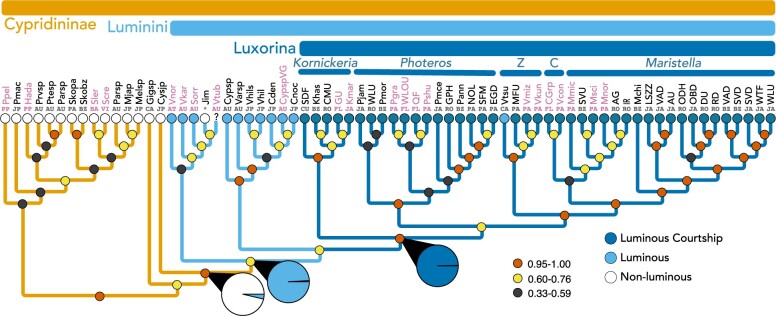
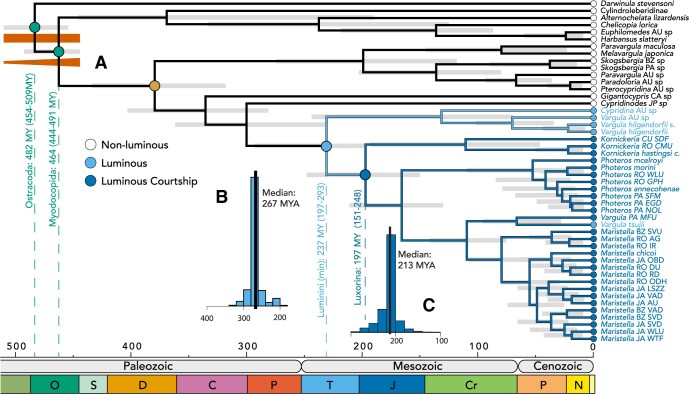
Although the diversity, beauty, and intricacy of sexually selected courtship displays command the attention of evolutionists, the longevity of these traits in deep time is poorly understood. Population-based theory suggests sexual selection could either lower or raise extinction risk, resulting in high or low persistence of lineages with sexually selected traits. Furthermore, empirical studies that directly estimate the longevity of sexually selected traits are uncommon. Sexually selected signals-including bioluminescent courtship-originated multiple times during evolution, allowing the empirical study of their longevity after careful phylogenetic and divergence time analyses. Here, we estimate the first transcriptome-based molecular phylogeny and divergence times of Cypridinidae. We report extreme longevity of bioluminescent courtship, a trait important in mate choice and probably under sexual selection. Our relaxed-clock estimates of divergence times coupled with stochastic character mapping show luminous courtship evolved only once in Cypridinidae-in a Sub-Tribe, we name Luxorina-at least 151 millions of years ago from cypridinid ancestors that used bioluminescence only in antipredator displays, defining a Tribe we name Luminini. This time-calibrated molecular phylogeny of cypridinids will serve as a foundation for integrative and comparative studies on the biochemistry, molecular evolution, courtship, diversification, and ecology of cypridinid bioluminescence. The persistence of luminous courtship for hundreds of millions of years suggests that sexual selection did not cause a rapid loss of associated traits, and that rates of speciation within the group exceeded extinction risk, which may contribute to the persistence of a diverse clade of signaling species. [Ancestral state reconstruction; Biodiversity; co-option; divergence time estimates; macroevolution; Ostracoda; phylogenomics; sexual selection.].
© The Author(s) 2022. Published by Oxford University Press on behalf of the Society of Systematic Biologists.
Figures
Similar articles
-
High Rates of Species Accumulation in Animals with Bioluminescent Courtship Displays.Curr Biol. 2016 Jul 25;26(14):1916-21. doi: 10.1016/j.cub.2016.05.043. Epub 2016 Jun 23. Curr Biol. 2016. PMID: 27345160
-
Laboratory culture of the California Sea Firefly Vargula tsujii (Ostracoda: Cypridinidae): Developing a model system for the evolution of marine bioluminescence.Sci Rep. 2020 Jun 26;10(1):10443. doi: 10.1038/s41598-020-67209-w. Sci Rep. 2020. PMID: 32591605 Free PMC article.
-
Phenotypic evolution shaped by current enzyme function in the bioluminescent courtship signals of sea fireflies.Proc Biol Sci. 2019 Jan 16;286(1894):20182621. doi: 10.1098/rspb.2018.2621. Proc Biol Sci. 2019. PMID: 30963873 Free PMC article.
-
Songs versus colours versus horns: what explains the diversity of sexually selected traits?Biol Rev Camb Philos Soc. 2020 Aug;95(4):847-864. doi: 10.1111/brv.12593. Epub 2020 Feb 24. Biol Rev Camb Philos Soc. 2020. PMID: 32092241
-
Glucocorticoids, male sexual signals, and mate choice by females: Implications for sexual selection.Gen Comp Endocrinol. 2020 Mar 1;288:113354. doi: 10.1016/j.ygcen.2019.113354. Epub 2019 Dec 9. Gen Comp Endocrinol. 2020. PMID: 31830474 Review.
Cited by
-
Similar enzymatic functions in distinct bioluminescence systems: evolutionary recruitment of sulfotransferases in ostracod light organs.Biol Lett. 2024 May;20(5):20230585. doi: 10.1098/rsbl.2023.0585. Epub 2024 May 15. Biol Lett. 2024. PMID: 38746983 Free PMC article.
-
Dataset from RNAseq analysis of differential gene expression among developmental stages of two non-marine ostracodes.Data Brief. 2024 Jan 17;53:110070. doi: 10.1016/j.dib.2024.110070. eCollection 2024 Apr. Data Brief. 2024. PMID: 38317728 Free PMC article.
-
A brittle star is born: Ontogeny of luminous capabilities in Amphiura filiformis.PLoS One. 2024 Mar 11;19(3):e0298185. doi: 10.1371/journal.pone.0298185. eCollection 2024. PLoS One. 2024. PMID: 38466680 Free PMC article.
-
Collective synchrony of mating signals modulated by ecological cues and social signals in bioluminescent sea fireflies.Proc Biol Sci. 2023 Nov 29;290(2011):20232311. doi: 10.1098/rspb.2023.2311. Epub 2023 Nov 29. Proc Biol Sci. 2023. PMID: 38018106 Free PMC article.
-
Major Revisions in Pancrustacean Phylogeny and Evidence of Sensitivity to Taxon Sampling.Mol Biol Evol. 2023 Aug 3;40(8):msad175. doi: 10.1093/molbev/msad175. Mol Biol Evol. 2023. PMID: 37552897 Free PMC article. Review.
References
-
- Alexandrou M.A., Swartz B.A., Matzke N.J., Oakley T.H.. 2013. Genome duplication and multiple evolutionary origins of complex migratory behavior in Salmonidae. Mol. Phylogenet. Evol. 69:514–523. - PubMed
-
- Andersson M.B. 1994. Sexual selection. Princeton (NJ): Princeton University Press.
-
- Baumgartner P.O., Rojas-Agramonte Y., Sandoval-Gutierrez M., Urbani F., García-Delgado D., Garban G., Pérez Rodríguez M.. 2013. Late Jurassic breakup of the Proto-Caribbean and circum-global circulation across Pangea. EGU General Assembly Conference Abstracts. 13408.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
