

The pre-exposure SARS-CoV-2-specific T cell repertoire determines the quality of the immune response to vaccination
- PMID: 35985324
- PMCID: PMC9372089
- DOI: 10.1016/j.immuni.2022.08.003
The pre-exposure SARS-CoV-2-specific T cell repertoire determines the quality of the immune response to vaccination
Abstract
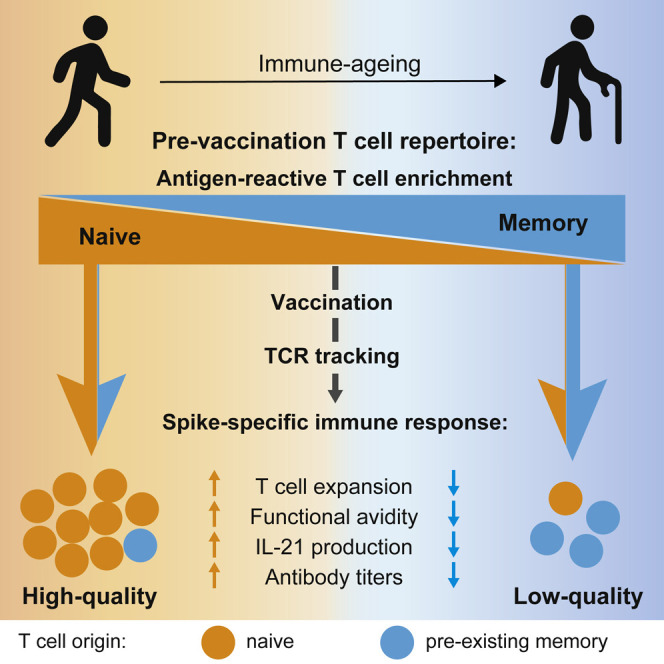
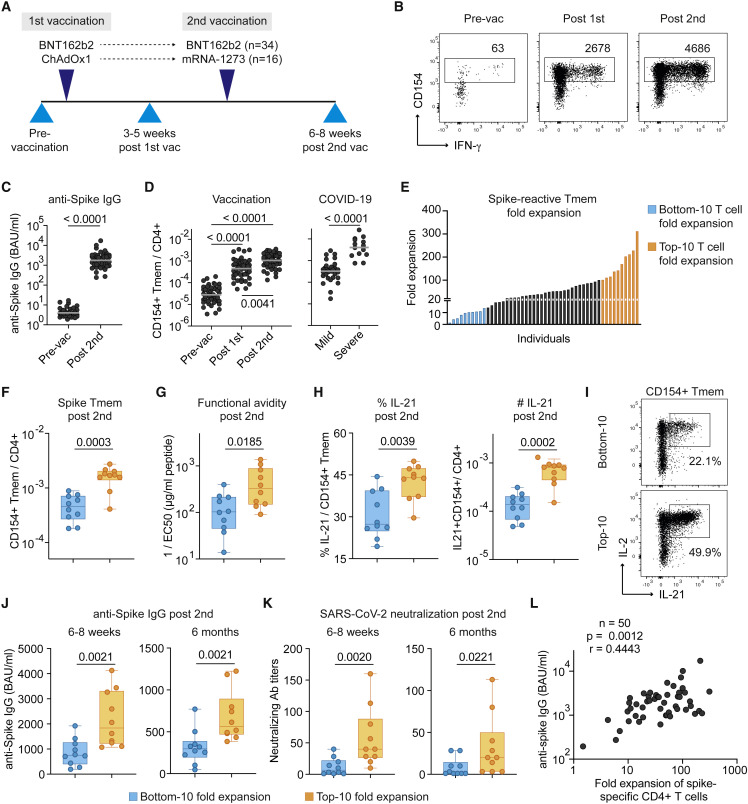
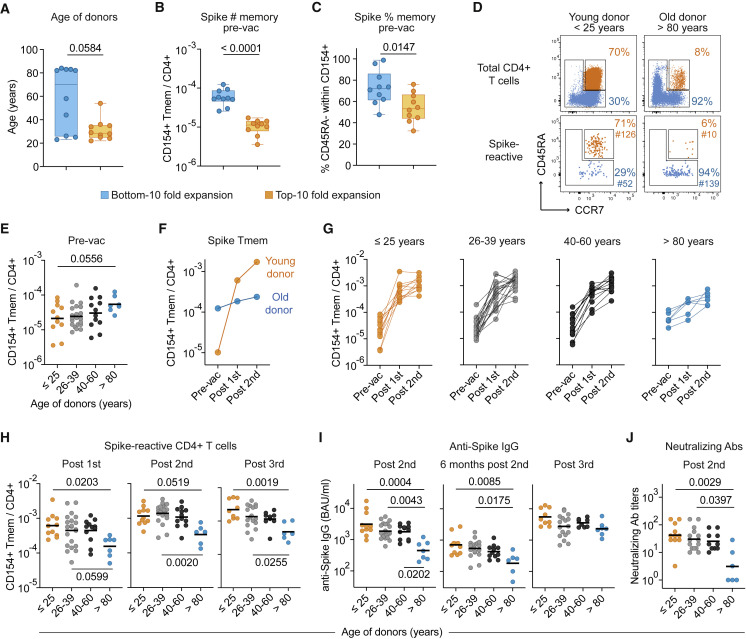
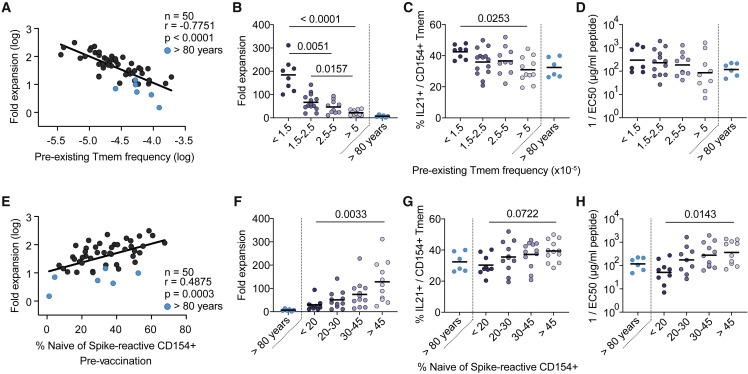
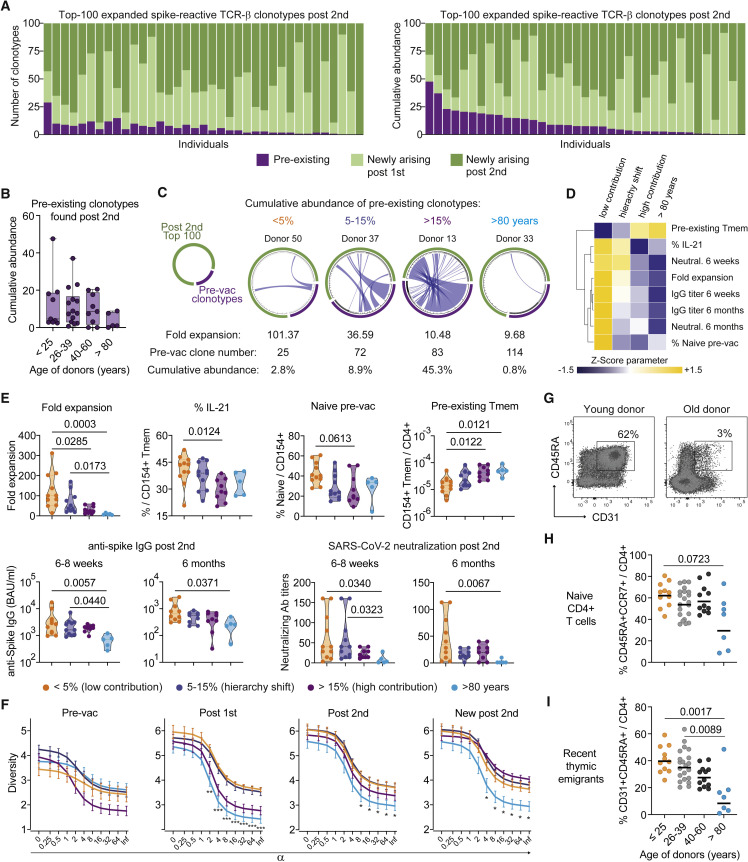
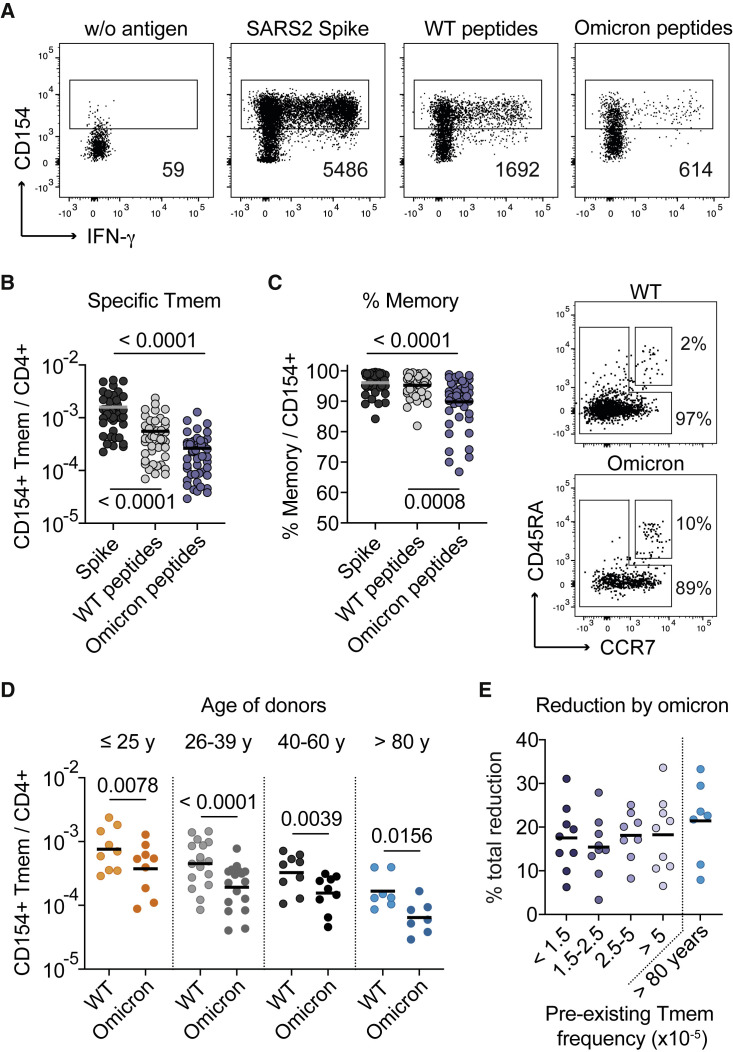
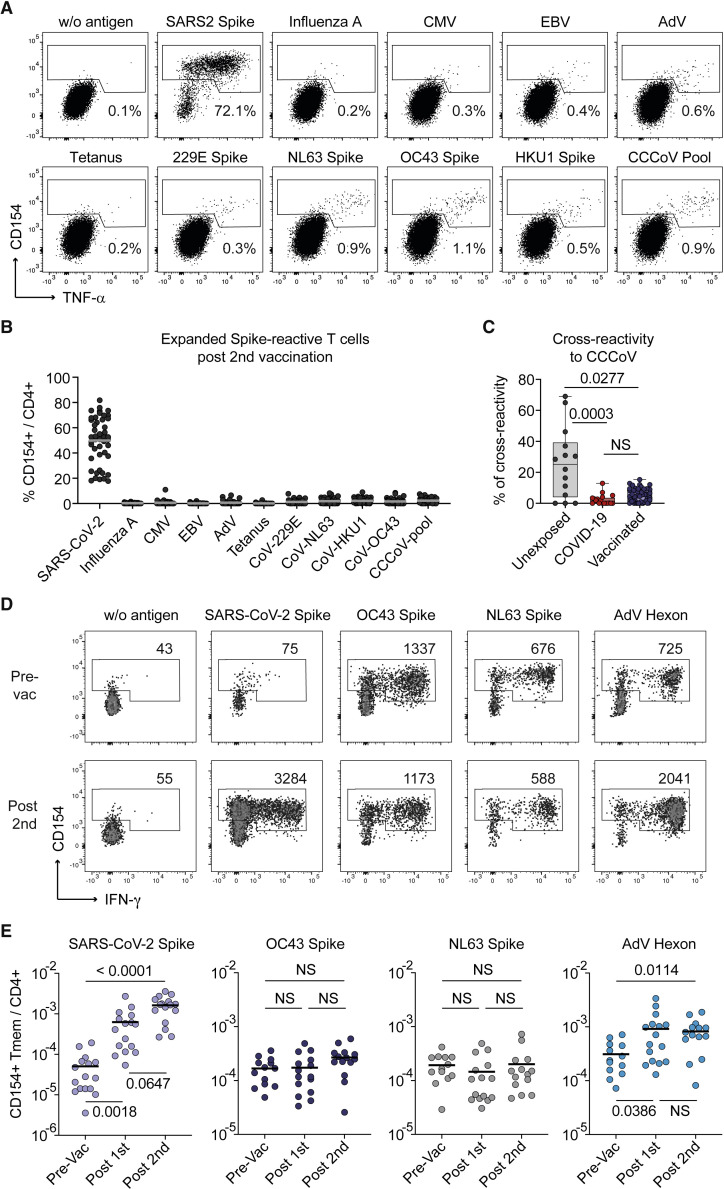
SARS-CoV-2 infection and vaccination generates enormous host-response heterogeneity and an age-dependent loss of immune-response quality. How the pre-exposure T cell repertoire contributes to this heterogeneity is poorly understood. We combined analysis of SARS-CoV-2-specific CD4+ T cells pre- and post-vaccination with longitudinal T cell receptor tracking. We identified strong pre-exposure T cell variability that correlated with subsequent immune-response quality and age. High-quality responses, defined by strong expansion of high-avidity spike-specific T cells, high interleukin-21 production, and specific immunoglobulin G, depended on an intact naive repertoire and exclusion of pre-existing memory T cells. In the elderly, T cell expansion from both compartments was severely compromised. Our results reveal that an intrinsic defect of the CD4+ T cell repertoire causes the age-dependent decline of immune-response quality against SARS-CoV-2 and highlight the need for alternative strategies to induce high-quality T cell responses against newly arising pathogens in the elderly.
Keywords: CD154; CD40L; COVID-19; SARS-CoV-2; TCR tracking; antigen-reactive T cell enrichment; antigen-specific CD4+ T cells; vaccination.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests E.R. is employee of Evotec SE. O.K. is employee of Peptides & elephants GmbH. O.C. reports grants or contracts from MSD, Pfizer; consulting fees from Biocon, Molecular Partners, Noxxon, PSI; honoraria for lectures from Abbott, Pfizer; participation on a data safety monitoring board or advisory board from Jannsen, PSI. P.K. reports grants or contracts from BMBF, B-FAST, and NAPKON of the Network University Medicine and the State of North Rhine-Westphalia; consulting fees from Ambu GmbH, Gilead Sciences, Noxxon N.V., Pfizer Pharma; honoraria for lectures from Ambu GmbH, Gilead Sciences, MSD Sharp & Dohme GmbH, Pfizer Pharma GmbH, Scilink Comunicación Científica SC; participation on an advisory board from Ambu GmbH, Gilead Sciences, Pfizer Pharma.
Figures
Similar articles
-
Persistent Maintenance of Intermediate Memory B Cells Following SARS-CoV-2 Infection and Vaccination Recall Response.J Virol. 2022 Aug 10;96(15):e0076022. doi: 10.1128/jvi.00760-22. Epub 2022 Jul 12. J Virol. 2022. PMID: 35862718 Free PMC article.
-
Antigen-Specific CD4+ T-Cell Activation in Primary Antibody Deficiency After BNT162b2 mRNA COVID-19 Vaccination.Front Immunol. 2022 Feb 14;13:827048. doi: 10.3389/fimmu.2022.827048. eCollection 2022. Front Immunol. 2022. PMID: 35237272 Free PMC article.
-
Architecture of the SARS-CoV-2-specific T cell repertoire.Front Immunol. 2023 Mar 20;14:1070077. doi: 10.3389/fimmu.2023.1070077. eCollection 2023. Front Immunol. 2023. PMID: 37020560 Free PMC article. Review.
-
Characterization of SARS-CoV-2-Specific Humoral and Cellular Immune Responses Induced by Inactivated COVID-19 Vaccines in a Real-World Setting.Front Immunol. 2021 Dec 22;12:802858. doi: 10.3389/fimmu.2021.802858. eCollection 2021. Front Immunol. 2021. PMID: 35003131 Free PMC article.
-
[COVID-19 Update].MMW Fortschr Med. 2023 Feb;165(2):58-61. doi: 10.1007/s15006-022-2232-x. MMW Fortschr Med. 2023. PMID: 36703066 Free PMC article. Review. German. No abstract available.
Cited by
-
Benefit delayed immunosenescence by regulating CD4+T cells: A promising therapeutic target for aging-related diseases.Aging Cell. 2024 Oct;23(10):e14317. doi: 10.1111/acel.14317. Epub 2024 Aug 18. Aging Cell. 2024. PMID: 39155409 Free PMC article. Review.
-
Nucleic acid vaccines: innovations, efficacy, and applications in at-risk populations.Front Immunol. 2025 May 14;16:1584876. doi: 10.3389/fimmu.2025.1584876. eCollection 2025. Front Immunol. 2025. PMID: 40438110 Free PMC article. Review.
-
Both Humoral and Cellular Immunity Limit the Ability of Live Attenuated Influenza Vaccines to Promote T Cell Responses.J Immunol. 2024 Jan 1;212(1):107-116. doi: 10.4049/jimmunol.2300343. J Immunol. 2024. PMID: 37982700 Free PMC article.
-
Antigen-specific T helper cells and cytokine profiles predict intensity and longevity of cellular and humoral responses to SARS-CoV-2 booster vaccination.Front Immunol. 2024 Aug 29;15:1423766. doi: 10.3389/fimmu.2024.1423766. eCollection 2024. Front Immunol. 2024. PMID: 39267758 Free PMC article.
-
Pre-existing T Cell Memory to Novel Pathogens.Immunohorizons. 2023 Jul 1;7(7):543-553. doi: 10.4049/immunohorizons.2200003. Immunohorizons. 2023. PMID: 37436166 Free PMC article. Review.
References
-
- Anderson E.J., Rouphael N.G., Widge A.T., Jackson L.A., Roberts P.C., Makhene M., Chappell J.D., Denison M.R., Stevens L.J., Pruijssers A.J., et al. Safety and immunogenicity of SARS-CoV-2 mRNA-1273 vaccine in older adults. N. Engl. J. Med. 2020;383:2427–2438. doi: 10.1056/NEJMoa2028436. - DOI - PMC - PubMed
-
- Bacher P., Hohnstein T., Beerbaum E., Röcker M., Blango M.G., Kaufmann S., Röhmel J., Eschenhagen P., Grehn C., Seidel K., et al. Human anti-fungal Th17 Immunity and pathology rely on cross-reactivity against candida albicans. Cell. 2019;176:1340–1355.e15. doi: 10.1016/j.cell.2019.01.041. - DOI - PubMed
-
- Bacher P., Rosati E., Esser D., Martini G.R., Saggau C., Schiminsky E., Dargvainiene J., Schröder I., Wieters I., Khodamoradi Y., et al. Low-Avidity CD4(+) T Cell responses to SARS-CoV-2 in unexposed individuals and humans with severe COVID-19. Immunity. 2020;53:1258–1271.e5. doi: 10.1016/j.immuni.2020.11.016. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
