

Faecalibaculum rodentium remodels retinoic acid signaling to govern eosinophil-dependent intestinal epithelial homeostasis
- PMID: 35985335
- PMCID: PMC9481734
- DOI: 10.1016/j.chom.2022.07.015
Faecalibaculum rodentium remodels retinoic acid signaling to govern eosinophil-dependent intestinal epithelial homeostasis
Abstract
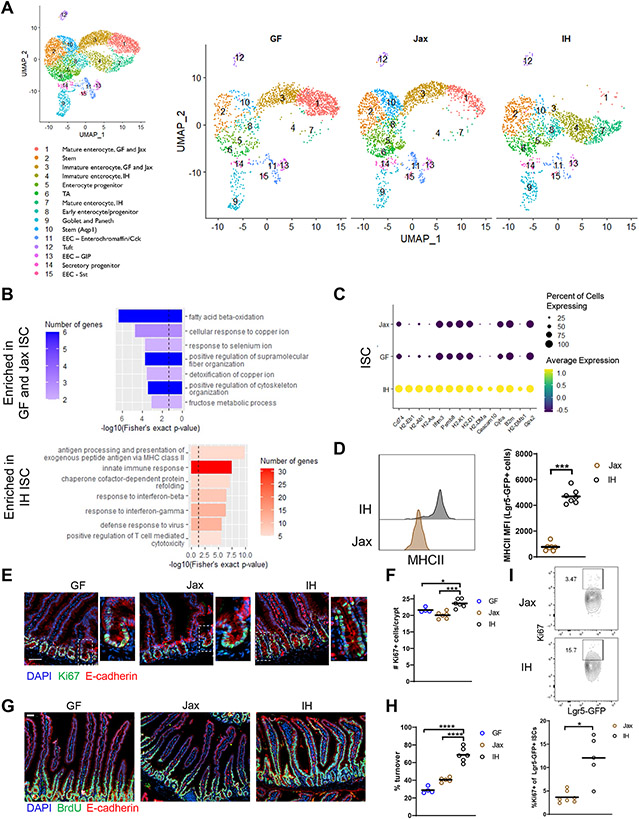
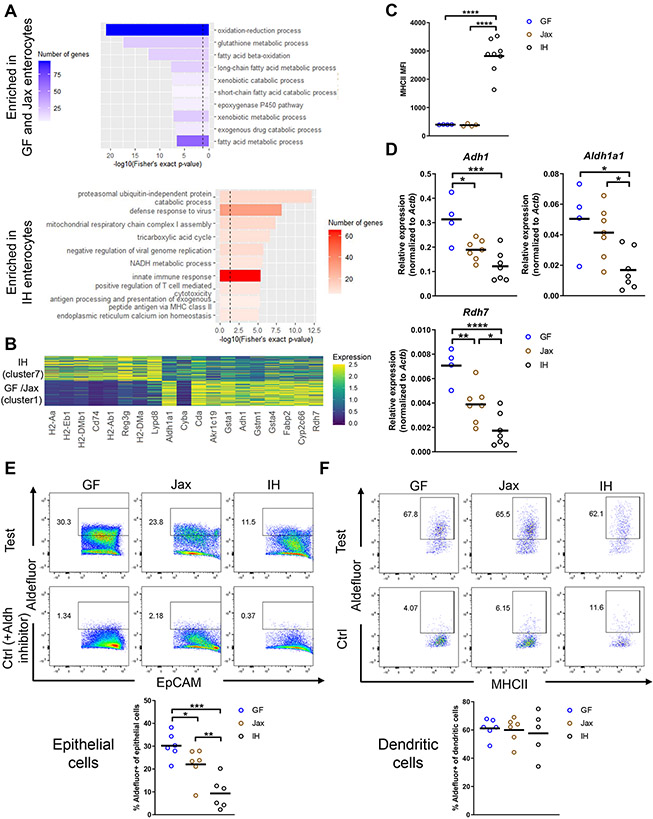
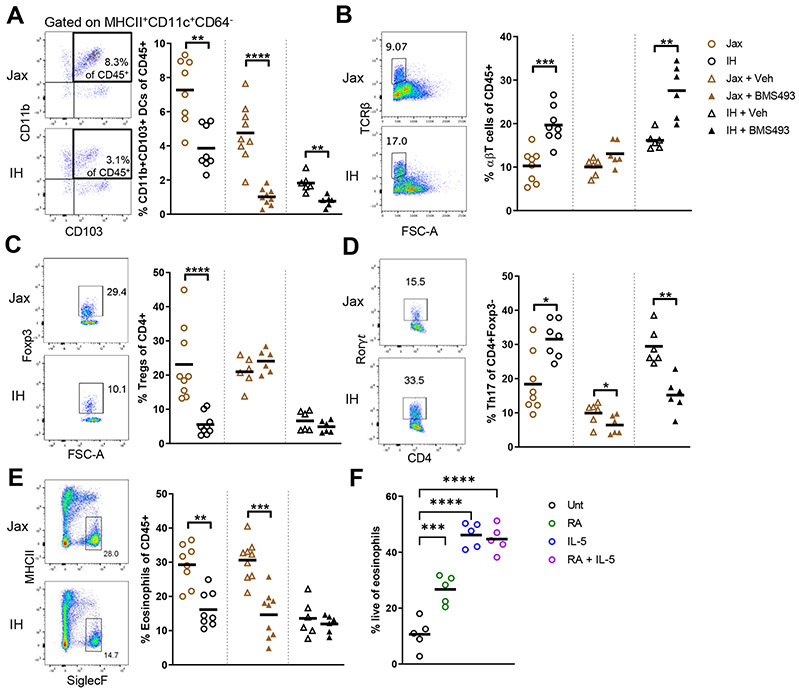
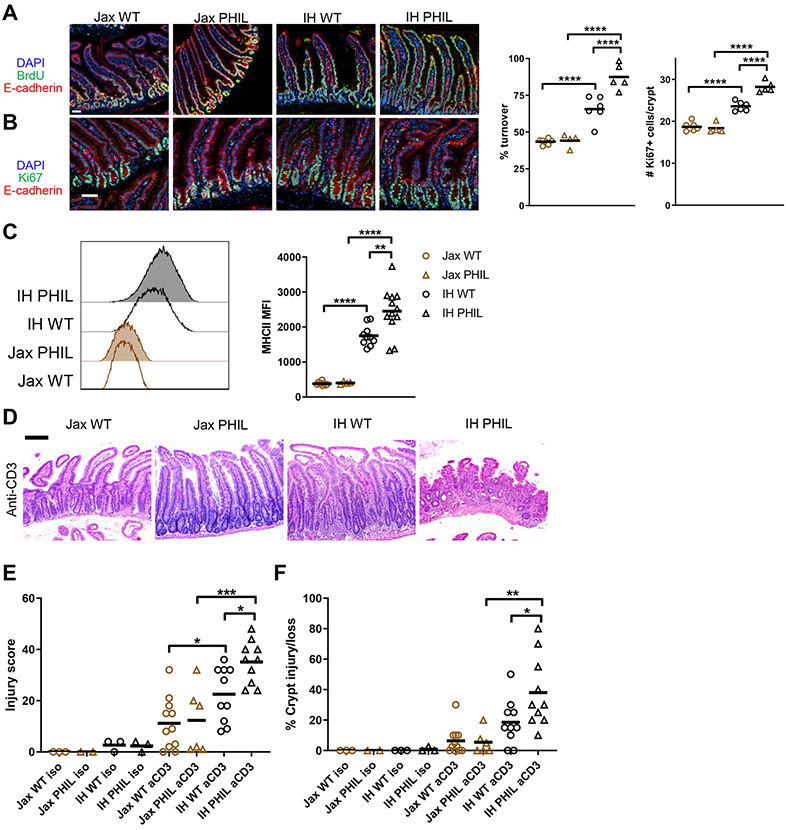
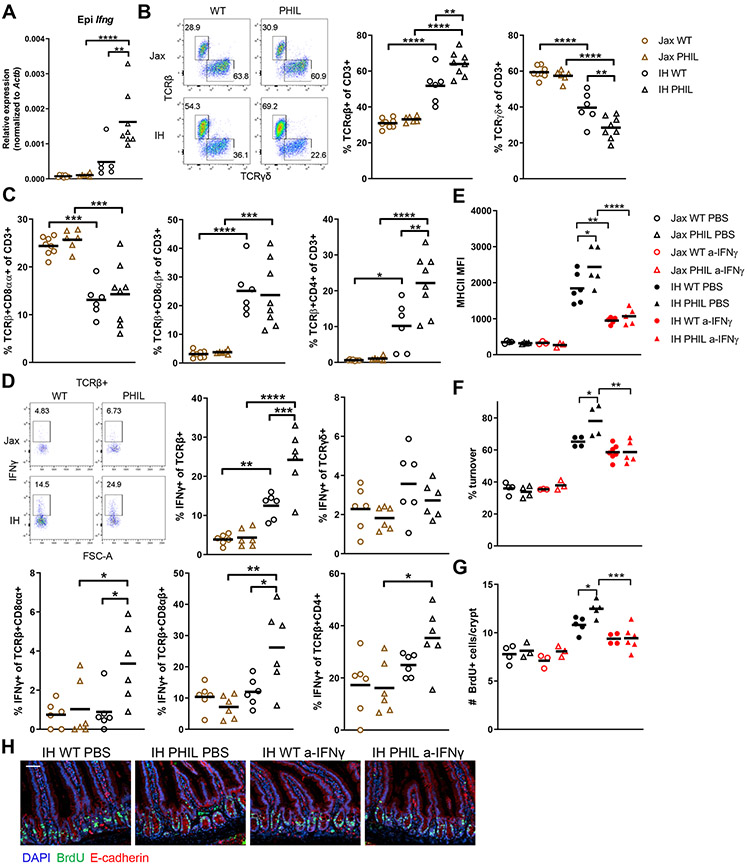
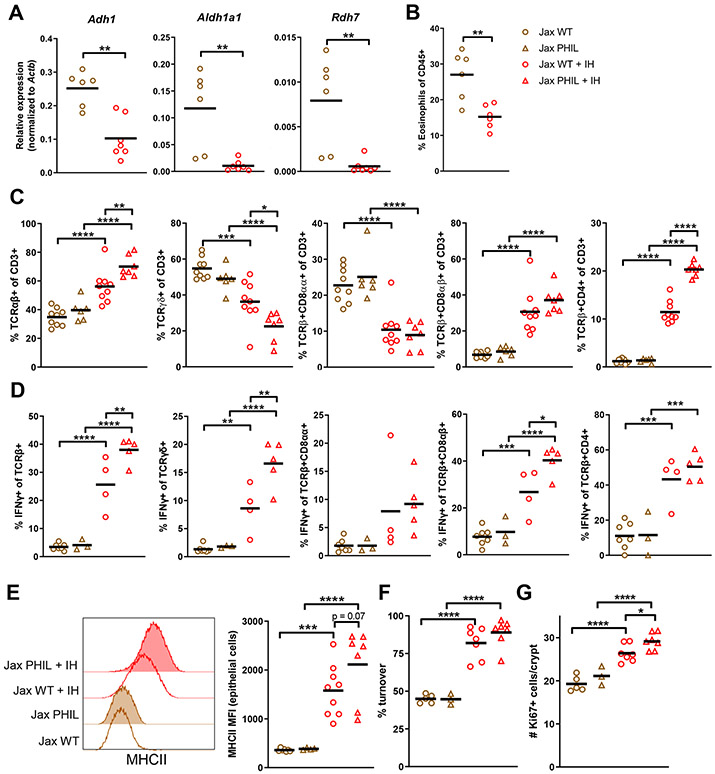
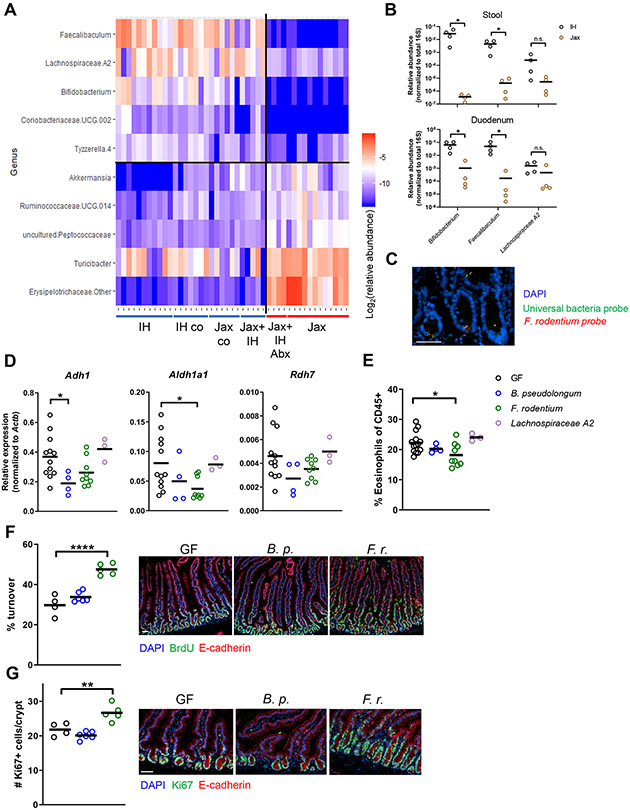
The intestinal epithelium plays critical roles in sensing and integrating dietary and microbial signals. How microbiota and intestinal epithelial cell (IEC) interactions regulate host physiology in the proximal small intestine, particularly the duodenum, is unclear. Using single-cell RNA sequencing of duodenal IECs under germ-free (GF) and different conventional microbiota compositions, we show that specific microbiota members alter epithelial homeostasis by increasing epithelial turnover rate, crypt proliferation, and major histocompatibility complex class II (MHCII) expression. Microbiome profiling identified Faecalibaculum rodentium as a key species involved in this regulation. F. rodentium decreases enterocyte expression of retinoic-acid-producing enzymes Adh1, Aldh1a1, and Rdh7, reducing retinoic acid signaling required to maintain certain intestinal eosinophil populations. Eosinophils suppress intraepithelial-lymphocyte-mediated production of interferon-γ that regulates epithelial cell function. Thus, we identify a retinoic acid-eosinophil-interferon-γ-dependent circuit by which the microbiota modulates duodenal epithelial homeostasis.
Keywords: Faecalibaculum rodentium; duodenum; eosinophil; interferon-γ; intestinal epithelial cell; microbiota; retinoic acid; small intestine.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests W.S.G. is on the SAB of Senda Biosciences, Freya Biosciences, and Artizan Biosciences, all outside the current work.
Figures
References
-
- Alexa A, Rahnenführer J, and Lengauer T (2006). Improved scoring of functional groups from gene expression data by decorrelating GO graph structure. Bioinformatics 22, 1600–1607. - PubMed
-
- Allaire JM, Crowley SM, Law HT, Chang S-Y, Ko H-J, and Vallance BA (2018). The Intestinal Epithelium: Central Coordinator of Mucosal Immunity. Trends Immunol. 39, 677–696. - PubMed
-
- Balmer JE, and Blomhoff R (2002). Gene expression regulation by retinoic acid. J. Lipid Res 43, 1773–1808. - PubMed
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
