

An expanded whole-cell model of E. coli links cellular physiology with mechanisms of growth rate control
- PMID: 35986058
- PMCID: PMC9391491
- DOI: 10.1038/s41540-022-00242-9
An expanded whole-cell model of E. coli links cellular physiology with mechanisms of growth rate control
Abstract
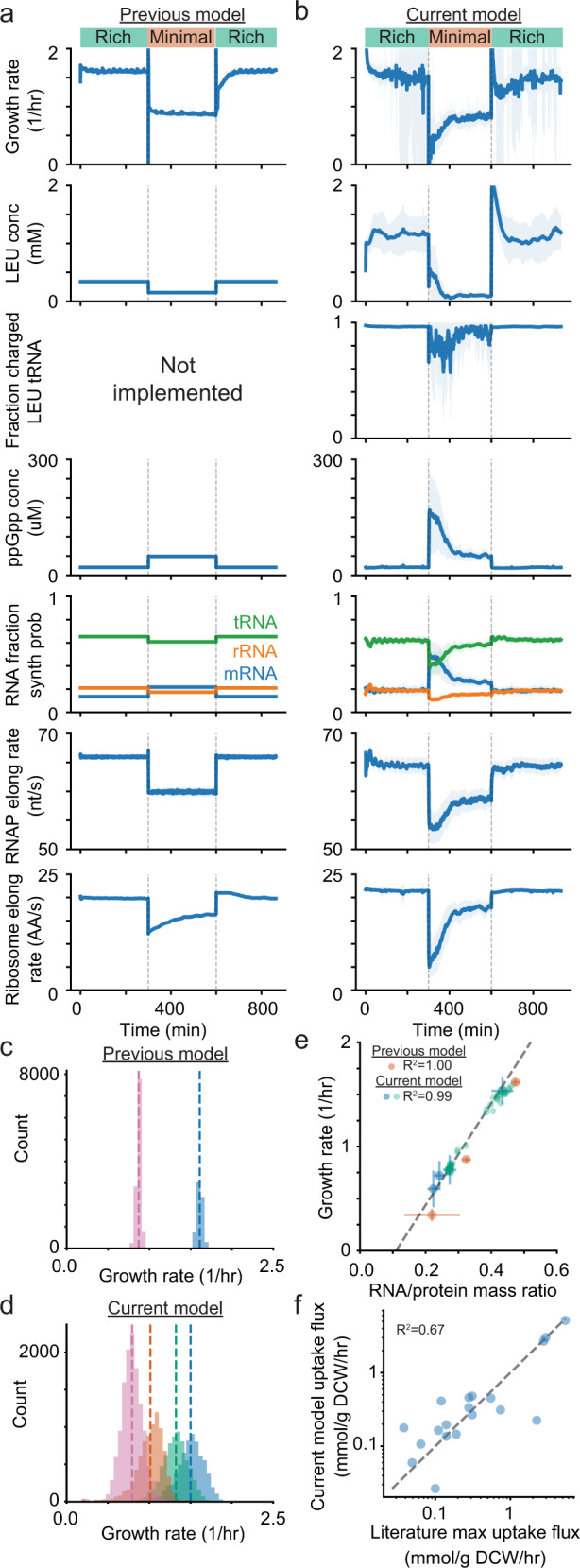
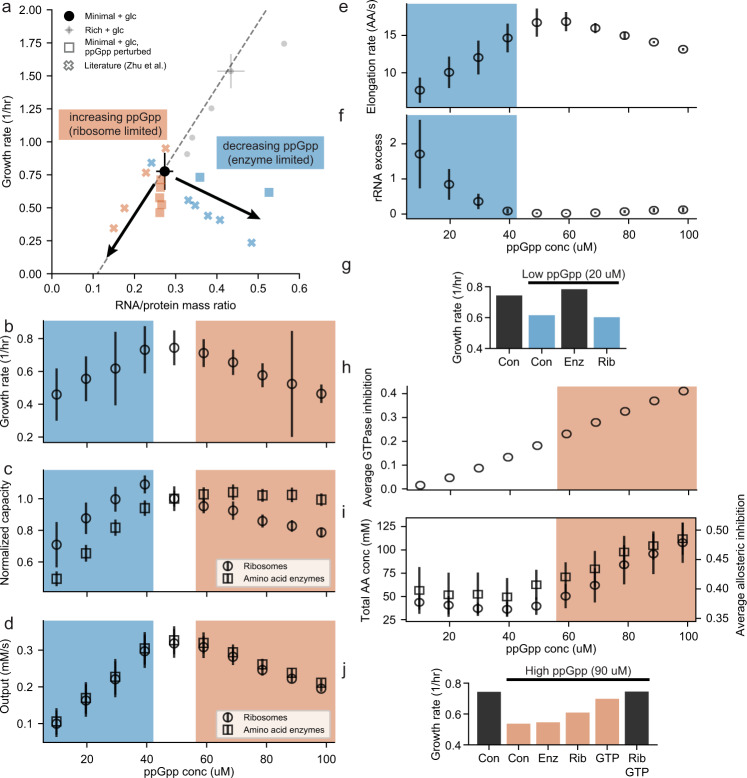
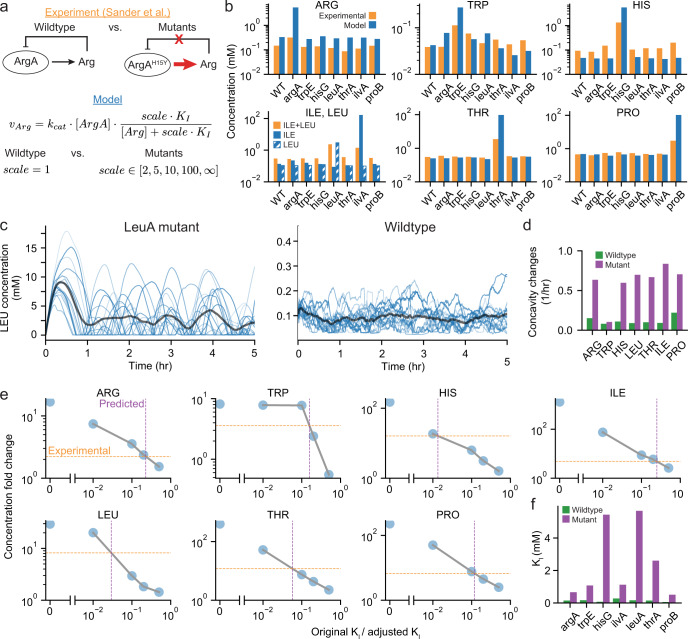
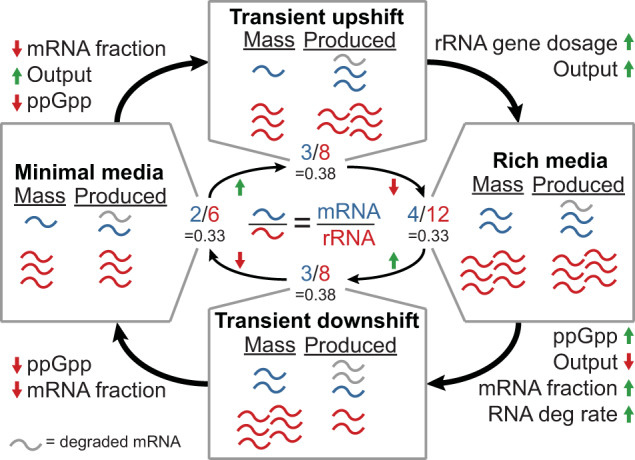
Growth and environmental responses are essential for living organisms to survive and adapt to constantly changing environments. In order to simulate new conditions and capture dynamic responses to environmental shifts in a developing whole-cell model of E. coli, we incorporated additional regulation, including dynamics of the global regulator guanosine tetraphosphate (ppGpp), along with dynamics of amino acid biosynthesis and translation. With the model, we show that under perturbed ppGpp conditions, small molecule feedback inhibition pathways, in addition to regulation of expression, play a role in ppGpp regulation of growth. We also found that simulations with dysregulated amino acid synthesis pathways provide average amino acid concentration predictions that are comparable to experimental results but on the single-cell level, concentrations unexpectedly show regular fluctuations. Additionally, during both an upshift and downshift in nutrient availability, the simulated cell responds similarly with a transient increase in the mRNA:rRNA ratio. This additional simulation functionality should support a variety of new applications and expansions of the E. coli Whole-Cell Modeling Project.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures


References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
