

Quantification of functional recovery in a larval zebrafish model of spinal cord injury
- PMID: 35986577
- PMCID: PMC10695274
- DOI: 10.1002/jnr.25118
Quantification of functional recovery in a larval zebrafish model of spinal cord injury
Abstract
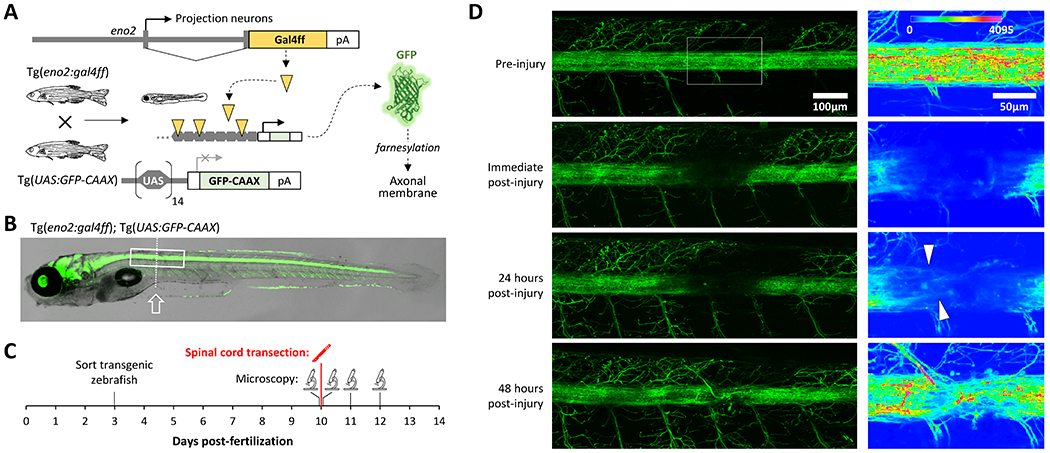
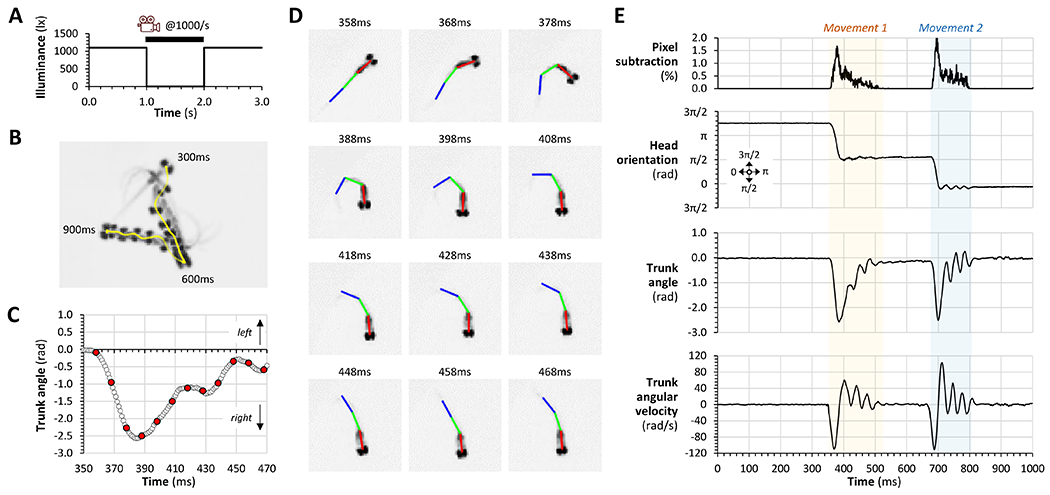
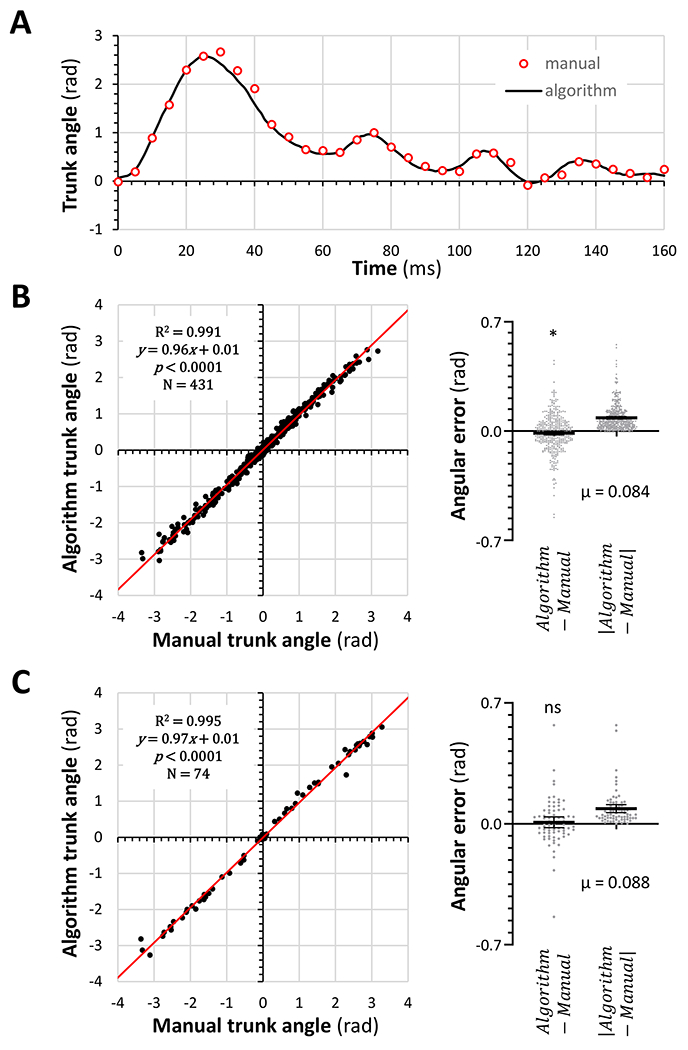
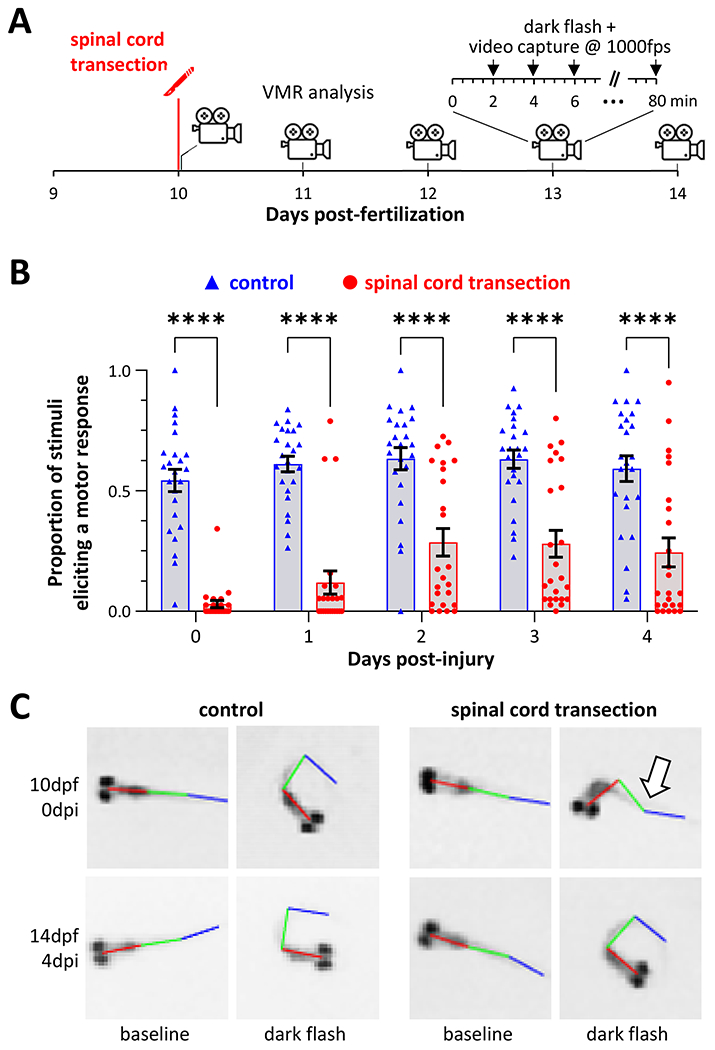
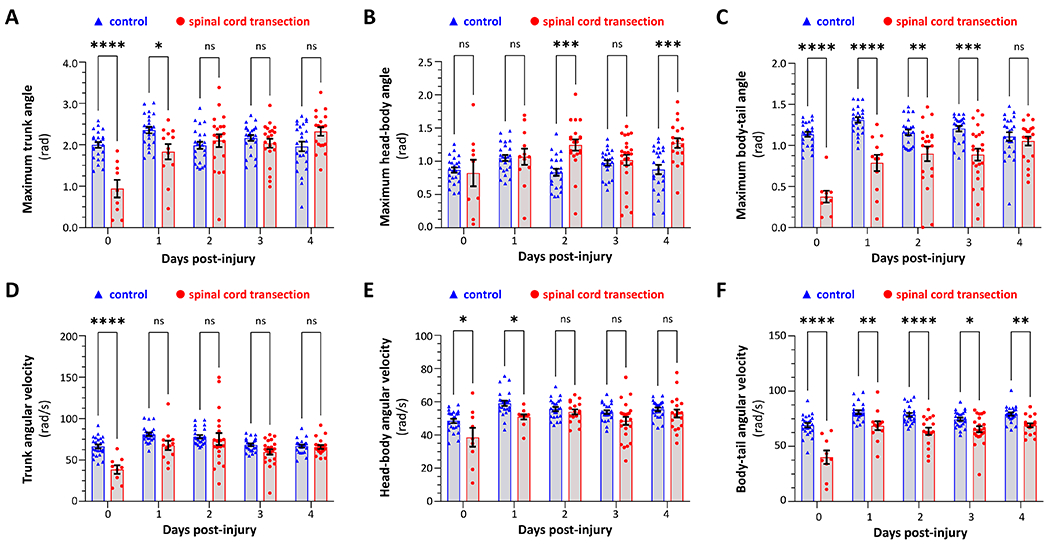
Human spinal cord injury (SCI) is characterized by permanent loss of damaged axons, resulting in chronic disability. In contrast, zebrafish can regenerate axonal projections following central nervous system injury and re-establish synaptic contacts with distant targets; elucidation of the underlying molecular events is an important goal with translational potential for improving outcomes in SCI patients. We generated transgenic zebrafish with GFP-labeled axons and transected their spinal cords at 10 days post-fertilization. Intravital confocal microscopy revealed robust axonal regeneration following the procedure, with abundant axons bridging the transection site by 48 h post-injury. In order to analyze neurological function in this model, we developed and validated new open-source software to measure zebrafish lateral trunk curvature during propulsive and turning movements at high temporal resolution. Immediately following spinal cord transection, axial movements were dramatically decreased caudal to the lesion site, but preserved rostral to the injury, suggesting the induction of motor paralysis below the transection level. Over the subsequent 96 h, the magnitude of movements caudal to the lesion recovered to baseline, but the rate of change of truncal curvature did not fully recover, suggesting incomplete restoration of caudal strength over this time course. Quantification of both morphological and functional recovery following SCI will be important for the analysis of axonal regeneration and downstream events necessary for restoration of motor function. An extensive array of genetic and pharmacological interventions can be deployed in the larval zebrafish model to investigate the underlying molecular mechanisms.
Keywords: axonal regeneration; high-speed macrovideography; intravital microscopy; machine vision; spinal paralysis; swimming kinematics.
© 2022 Wiley Periodicals LLC.
Conflict of interest statement
Conflict of interest statement:
The authors have no conflicts of interest.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
