

Warburg-like metabolic transformation underlies neuronal degeneration in sporadic Alzheimer's disease
- PMID: 35987203
- PMCID: PMC9458870
- DOI: 10.1016/j.cmet.2022.07.014
Warburg-like metabolic transformation underlies neuronal degeneration in sporadic Alzheimer's disease
Abstract
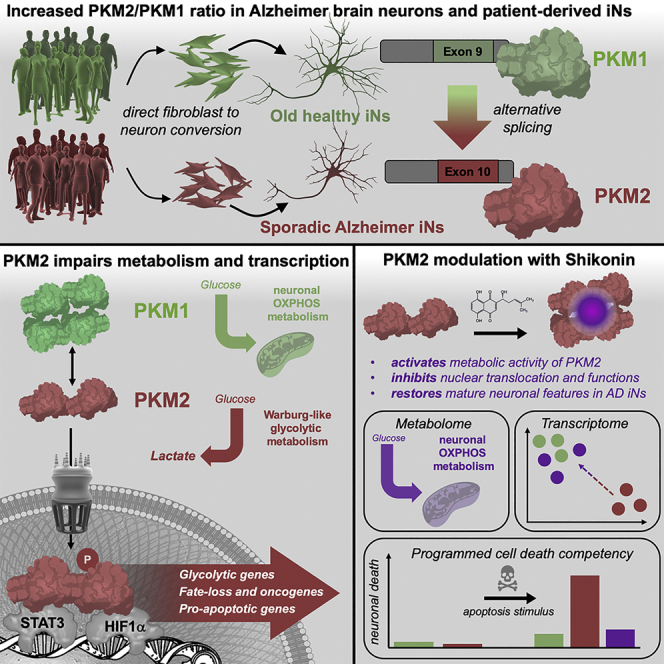
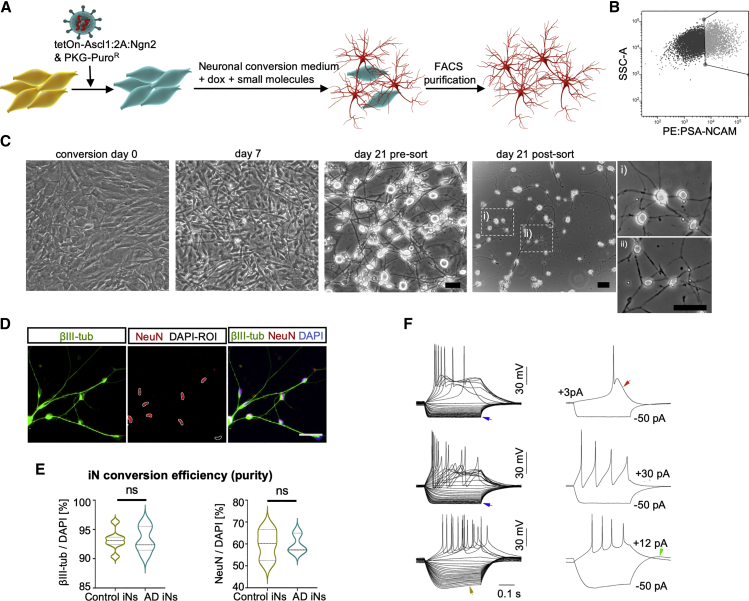
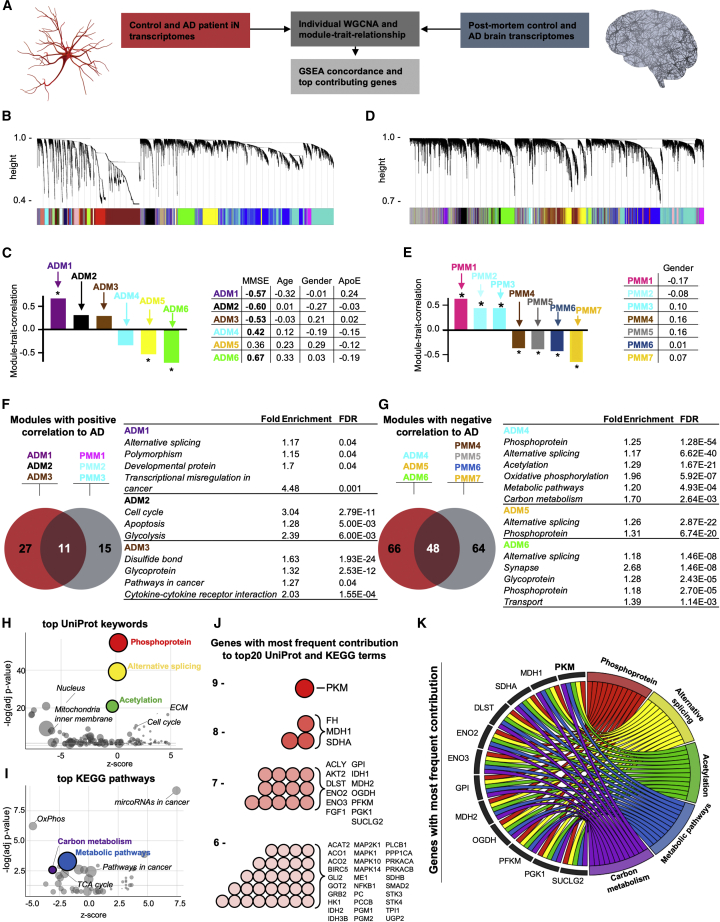
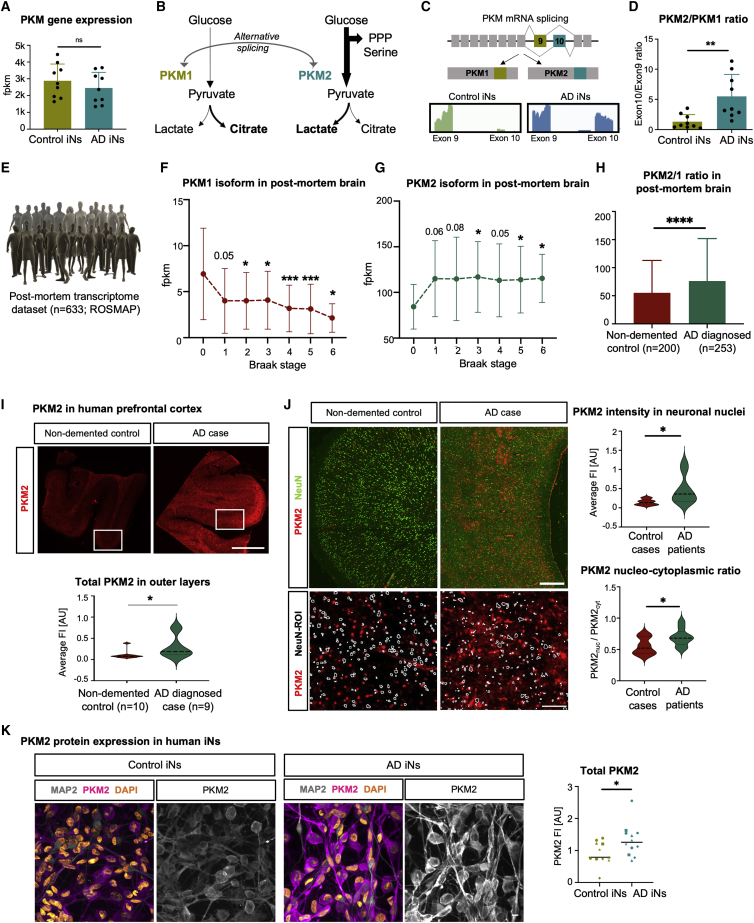
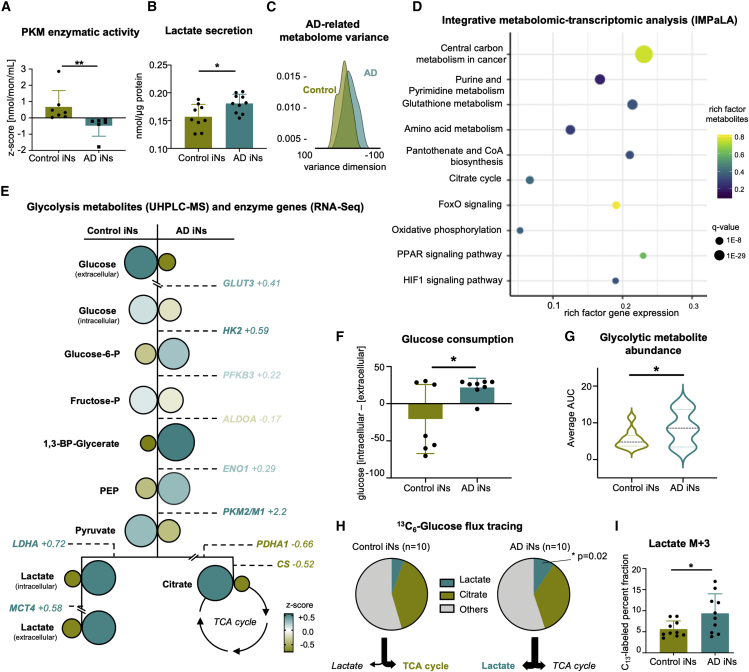
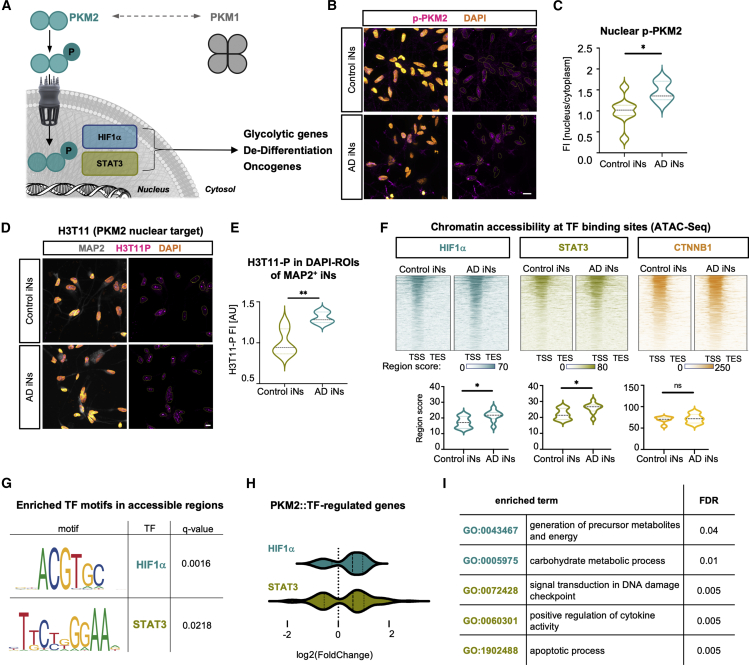
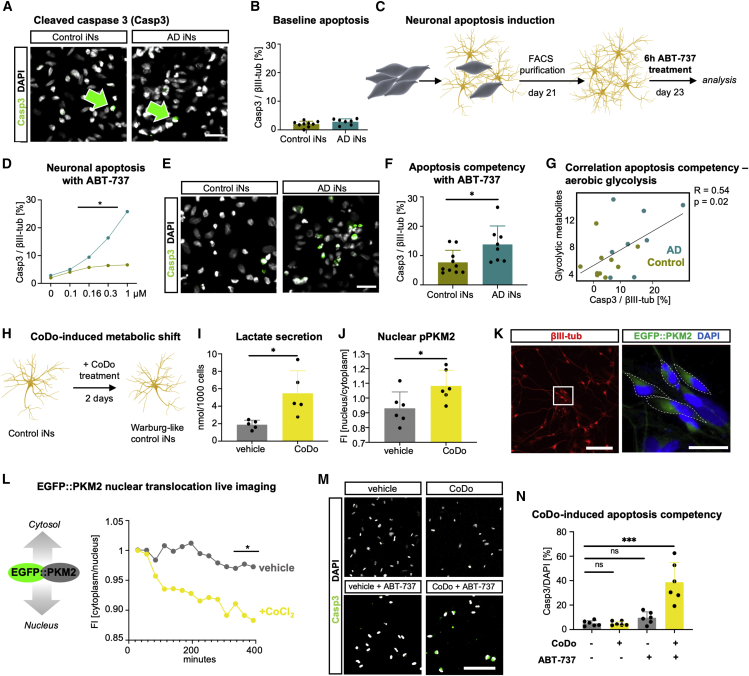
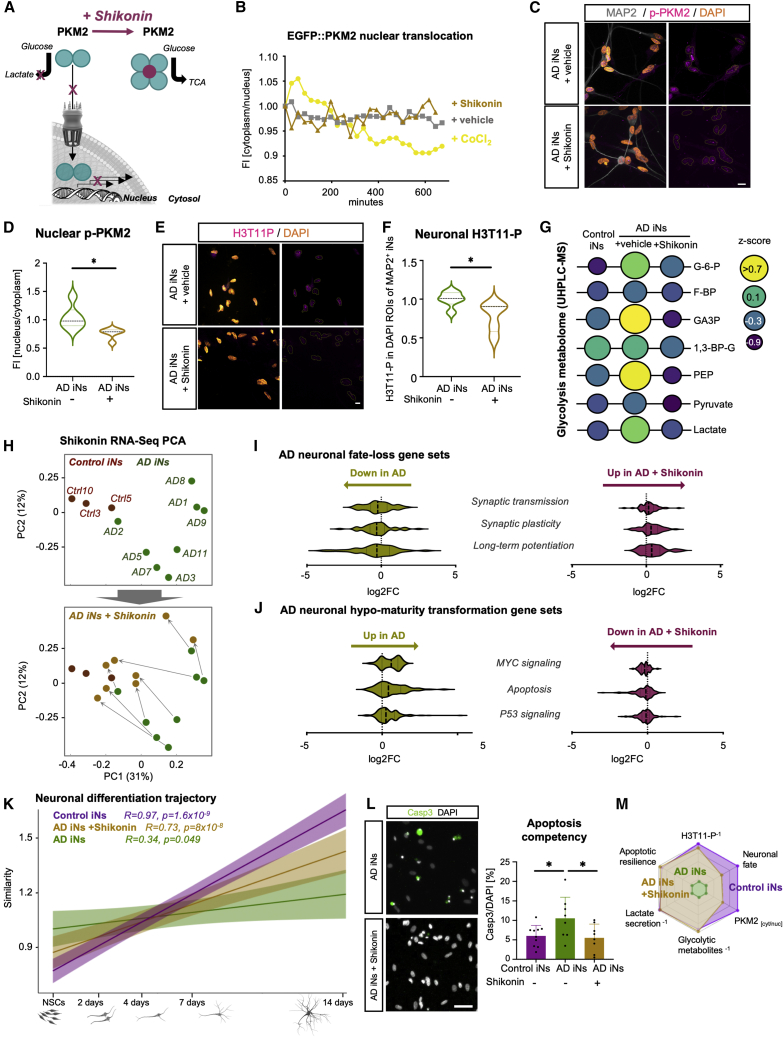
The drivers of sporadic Alzheimer's disease (AD) remain incompletely understood. Utilizing directly converted induced neurons (iNs) from AD-patient-derived fibroblasts, we identified a metabolic switch to aerobic glycolysis in AD iNs. Pathological isoform switching of the glycolytic enzyme pyruvate kinase M (PKM) toward the cancer-associated PKM2 isoform conferred metabolic and transcriptional changes in AD iNs. These alterations occurred via PKM2's lack of metabolic activity and via nuclear translocation and association with STAT3 and HIF1α to promote neuronal fate loss and vulnerability. Chemical modulation of PKM2 prevented nuclear translocation, restored a mature neuronal metabolism, reversed AD-specific gene expression changes, and re-activated neuronal resilience against cell death.
Keywords: Alzheimer's disease; WGCNA; Warburg effect; cancer; direct conversion; induced neurons; metabolomics; pyruvate kinase M; reprogramming.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Walking the high wire: How neurons maintain stability in the crossline of neurodegeneration.Cell Metab. 2022 Sep 6;34(9):1227-1229. doi: 10.1016/j.cmet.2022.08.013. Cell Metab. 2022. PMID: 36070677
References
-
- Arendt T., Holzer M., Stöbe A., Gärtner U., Lüth H.J., Brückner M.K., Ueberham U. Activated mitogenic signaling induces a process of dedifferentiation in Alzheimer’s disease that eventually results in cell death. Ann. N. Y. Acad. Sci. 2000;920:249–255. doi: 10.1111/j.1749-6632.2000.tb06931.x. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
