

The key molecular pattern BxCDP1 of Bursaphelenchus xylophilus induces plant immunity and enhances plant defense response via two small peptide regions
- PMID: 35991456
- PMCID: PMC9382027
- DOI: 10.3389/fpls.2022.937473
The key molecular pattern BxCDP1 of Bursaphelenchus xylophilus induces plant immunity and enhances plant defense response via two small peptide regions
Abstract
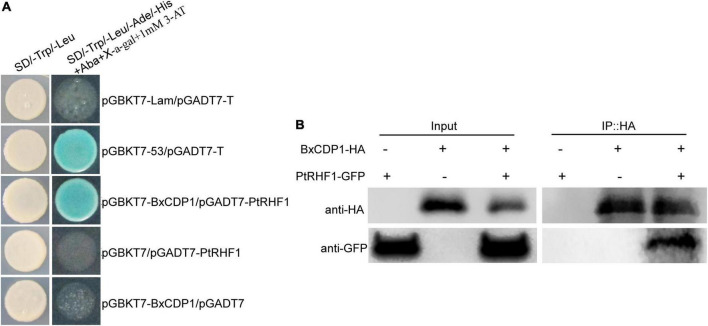
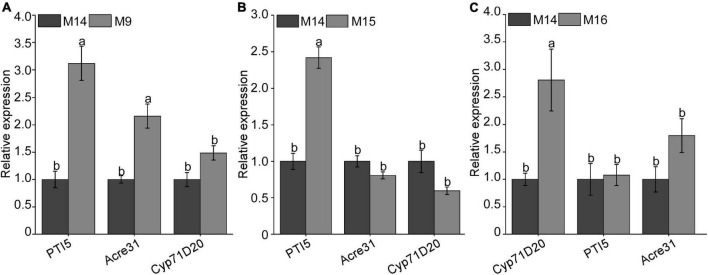
The migratory plant-parasitic nematode Bursaphelenchus xylophilus is the pathogen of the pine wilt disease (PWD), causing serious damage to pine forests in China. During the process of plant resistance to multiple pathogens, plant immunity plays a key role. In this current study, the pathogen-associated molecular pattern (PAMP) BxCDP1 in B. xylophilus has been identified, but the host target protein of BxCDP1 and its key amino acid region inducing the plant immunity have yet to be elucidated. We found that BxCDP1 could trigger superoxide production, H2O2 production, and callose deposits. A RING-H2 finger protein 1 (RHF1) of Pinus thunbergii was screened and characterized as a target protein of BxCDP1 by yeast two-hybrid and co-immunoprecipitation (Co-IP). Moreover, two peptides (namely M9 and M16) proved to be key regions of BxCDP1 to induce PAMP-triggered immunity (PTI) in Nicotiana benthamiana, which also induced the expression of pathogenesis-related (PR) genes (PtPR-3, PtPR-4, and PtPR-5) in P. thunbergii and enhanced the resistance of the host to B. xylophilus. These results indicate that BxCDP1 plays a critical role in the interaction between B. xylophilus and P. thunbergii, and both peptides M9 and M16 have the potential to be developed and utilized as immune inducers of pines against B. xylophilus in future.
Keywords: Bursaphelenchus xylophilus; Pinus thunbergii; immunity induction; key peptides; protein interaction.
Copyright © 2022 Hu, Wu, Wen, Ye, Qiu, Rui and Zhang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
-
- Chen J. S., Lin B. R., Huang Q. L., Hu L. L., Zhuo K., Liao J. L. (2017). A novel Meloidogyne graminicola effector, MgGPP, is secreted into host cells and undergoes glycosylation in concert with proteolysis to suppress plant defenses and promote parasitism. PLoS Pathog. 13:e1006301. 10.1371/journal.ppat.1006301 - DOI - PMC - PubMed
-
- Chisholm S. T., Coaker G., Day B., Staskawicz B. J. (2006). Host-microbe interactions: Shaping the evolution of the plant immune response. Cell 124 803–814. - PubMed
-
- Doke N. (1983). Involvement of superoxide anion generation in the hypersensitive response of potato tuber tissues to infection with an incompatible race of Phytophthora infestans and to the hyphal wall components. Physiol. Phant Pathol. 23 345–357.
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials