

Disrupting interaction between miR-132 and Mmp9 3'UTR improves synaptic plasticity and memory in mice
- PMID: 35992198
- PMCID: PMC9389266
- DOI: 10.3389/fnmol.2022.924534
Disrupting interaction between miR-132 and Mmp9 3'UTR improves synaptic plasticity and memory in mice
Abstract
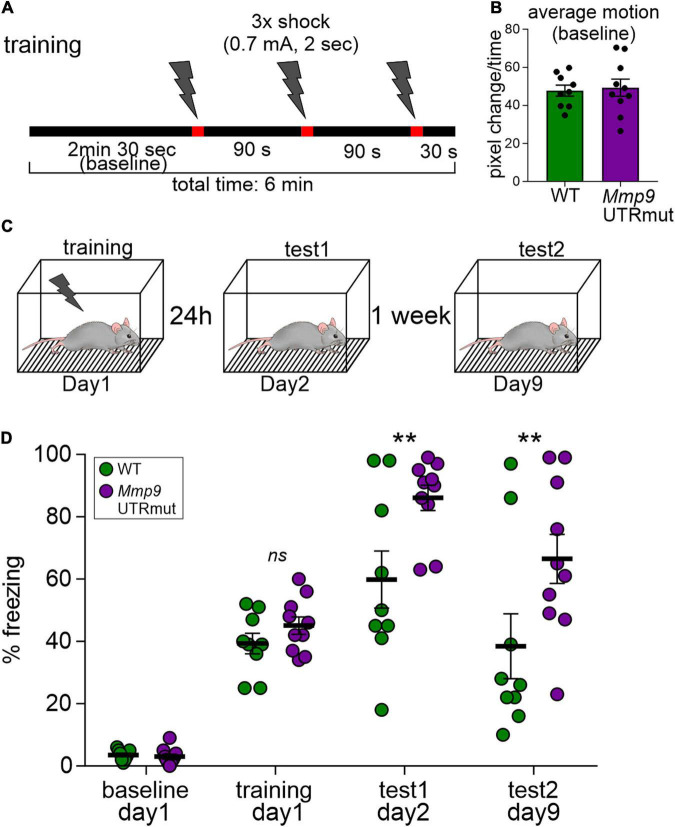
As microRNAs have emerged to be important regulators of molecular events occurring at the synapses, the new questions about their regulatory effect on the behavior have araised. In the present study, we show for the first time that the dysregulated specific targeting of miR132 to Mmp9 mRNA in the mouse brain results in the increased level of Mmp9 protein, which affects synaptic plasticity and has an effect on memory formation. Our data points at the importance of complex and precise regulation of the Mmp9 level by miR132 in the brain.
Keywords: Mmp9; behavior; brain; miR132; microRNA; synaptic plasticity.
Copyright © 2022 Kuzniewska, Rejmak, Nowacka, Ziółkowska, Milek, Magnowska, Gruchota, Gewartowska, Borsuk, Salamian, Dziembowski, Radwanska and Dziembowska.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures




References
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous