

Small molecule MMRi62 targets MDM4 for degradation and induces leukemic cell apoptosis regardless of p53 status
- PMID: 35992795
- PMCID: PMC9389462
- DOI: 10.3389/fonc.2022.933446
Small molecule MMRi62 targets MDM4 for degradation and induces leukemic cell apoptosis regardless of p53 status
Abstract
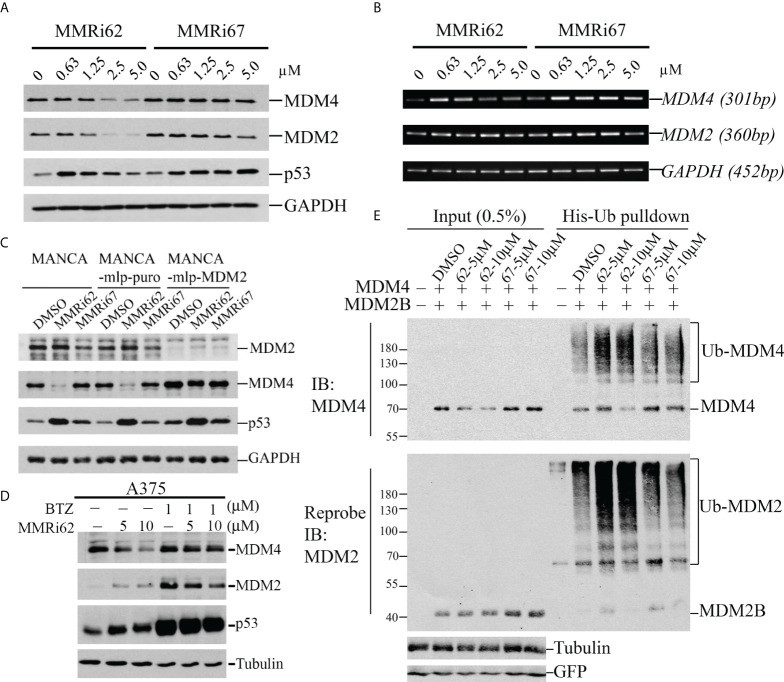
MDM2 and MDM4 proteins are key negative regulators of tumor suppressor p53. MDM2 and MDM4 interact via their RING domains and form a heterodimer polyubiquitin E3 ligase essential for p53 degradation. MDM4 also forms heterodimer E3 ligases with MDM2 isoforms that lack p53-binding domains, which regulate p53 and MDM4 stability. We are working to identify small-molecule inhibitors targeting the RING domain of MDM2-MDM4 (MMRi) that can inactivate the total oncogenic activity of MDM2-MDM4 heterodimers. Here, we describe the identification and characterization of MMRi62 as an MDM4-degrader and apoptosis inducer in leukemia cells. Biochemically, in our experiments, MMRi62 bound to preformed RING domain heterodimers altered the substrate preference toward MDM4 ubiquitination and promoted MDM2-dependent MDM4 degradation in cells. This MDM4-degrader activity of MMRi62 was found to be associated with potent apoptosis induction in leukemia cells. Interestingly, MMRi62 effectively induced apoptosis in p53 mutant, multidrug-resistant leukemia cells and patient samples in addition to p53 wild-type cells. In contrast, MMRi67 as a RING heterodimer disruptor and an enzymatic inhibitor of the MDM2-MDM4 E3 complex lacked MDM4-degrader activity and failed to induce apoptosis in these cells. In summary, this study identifies MMRi62 as a novel MDM2-MDM4-targeting agent and suggests that small molecules capable of promoting MDM4 degradation may be a viable new approach to killing leukemia cells bearing non-functional p53 by apoptosis.
Keywords: E3 ligase; MDM2; MDM4; apoptosis; degradation; leukemia; p53; ubiquitination.
Copyright © 2022 Lama, Xu, Galster, Querol-García, Portwood, Mavis, Ruiz, Martin, Wu, Giorgi, Bargonetti, Wang, Hernandez-Ilizaliturri, Koudelka, Chemler, Muñoz and Wang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





References
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous