

Differences in the Second Coordination Sphere Tailor the Substrate Specificity and Reactivity of Thiol Dioxygenases
- PMID: 35994511
- PMCID: PMC9583696
- DOI: 10.1021/acs.accounts.2c00359
Differences in the Second Coordination Sphere Tailor the Substrate Specificity and Reactivity of Thiol Dioxygenases
Abstract
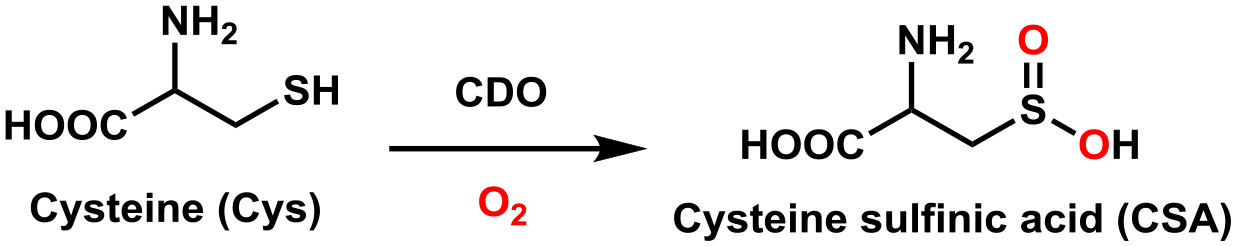
In recent years, considerable progress has been made toward elucidating the geometric and electronic structures of thiol dioxygenases (TDOs). TDOs catalyze the conversion of substrates with a sulfhydryl group to their sulfinic acid derivatives via the addition of both oxygen atoms from molecular oxygen. All TDOs discovered to date belong to the family of cupin-type mononuclear nonheme Fe(II)-dependent metalloenzymes. While most members of this enzyme family bind the Fe cofactor by two histidines and one carboxylate side chain (2-His-1-carboxylate) to provide a monoanionic binding motif, TDOs feature a neutral three histidine (3-His) facial triad. In this Account, we present a bioinformatics analysis and multiple sequence alignment that highlight the significance of the secondary coordination sphere in tailoring the substrate specificity and reactivity among the different TDOs. These insights provide the framework within which important structural and functional features of the distinct TDOs are discussed.The best studied TDO is cysteine dioxygenase (CDO), which catalyzes the conversion of cysteine to cysteine sulfinic acid in both eukaryotes and prokaryotes. Crystal structures of resting and substrate-bound mammalian CDOs revealed two surprising structural motifs in the first- and second coordination spheres of the Fe center. The first is the presence of the abovementioned neutral 3-His facial triad that coordinates the Fe ion. The second is the existence of a covalent cross-link between the sulfur of Cys93 and an ortho carbon of Tyr157 (mouse CDO numbering scheme). While the exact role of this cross-link remains incompletely understood, various studies established that it is needed for proper substrate Cys positioning and gating solvent access to the active site. Intriguingly, bacterial CDOs lack the Cys-Tyr cross-link; yet, they are as active as cross-linked eukaryotic CDOs.The other known mammalian TDO is cysteamine dioxygenase (ADO). Initially, it was believed that ADO solely catalyzes the oxidation of cysteamine to hypotaurine. However, it has recently been shown that ADO additionally oxidizes N-terminal cysteine (Nt-Cys) peptides, which indicates that ADO may play a much more significant role in mammalian physiology than was originally anticipated. Though predicted on the basis of sequence alignment, site-directed mutagenesis, and spectroscopic studies, it was not until last year that two crystal structures, one of wild-type mouse ADO (solved by us) and the other of a variant of nickel-substituted human ADO, finally provided direct evidence that this enzyme also features a 3-His facial triad. These structures additionally revealed several features that are unique to ADO, including a putative cosubstrate O2 access tunnel that is lined by two Cys residues. Disulfide formation under conditions of high O2 levels may serve as a gating mechanism to prevent ADO from depleting organisms of Nt-Cys-containing molecules.The combination of kinetic and spectroscopic studies in conjunction with structural characterizations of TDOs has furthered our understanding of enzymatic sulfhydryl substrate regulation. In this article, we take advantage of the fact that the ADO X-ray crystal structures provided the final piece needed to compare and contrast key features of TDOs, an essential family of metalloenzymes found across all kingdoms of life.
Conflict of interest statement
The authors declare no competing financial interest.
Figures












Similar articles
-
Non-standard amino acid incorporation into thiol dioxygenases.Methods Enzymol. 2024;703:121-145. doi: 10.1016/bs.mie.2024.05.022. Epub 2024 Jun 15. Methods Enzymol. 2024. PMID: 39260993 Free PMC article.
-
Spectroscopic investigation of iron(III) cysteamine dioxygenase in the presence of substrate (analogs): implications for the nature of substrate-bound reaction intermediates.J Biol Inorg Chem. 2021 Dec;26(8):947-955. doi: 10.1007/s00775-021-01904-5. Epub 2021 Sep 27. J Biol Inorg Chem. 2021. PMID: 34580769 Free PMC article.
-
Spectroscopic analysis of the mammalian enzyme cysteine dioxygenase.Methods Enzymol. 2023;682:101-135. doi: 10.1016/bs.mie.2023.01.002. Epub 2023 Feb 15. Methods Enzymol. 2023. PMID: 36948699 Free PMC article.
-
Thiol dioxygenases: unique families of cupin proteins.Amino Acids. 2011 Jun;41(1):91-102. doi: 10.1007/s00726-010-0518-2. Epub 2010 Mar 1. Amino Acids. 2011. PMID: 20195658 Free PMC article. Review.
-
Emerging roles for thiol dioxygenases as oxygen sensors.FEBS J. 2022 Sep;289(18):5426-5439. doi: 10.1111/febs.16147. Epub 2021 Aug 27. FEBS J. 2022. PMID: 34346181 Review.
Cited by
-
Cyanide replaces substrate in obligate-ordered addition of nitric oxide to the non-heme mononuclear iron AvMDO active site.J Biol Inorg Chem. 2023 Apr;28(3):285-299. doi: 10.1007/s00775-023-01990-7. Epub 2023 Feb 21. J Biol Inorg Chem. 2023. PMID: 36809458 Free PMC article.
-
Influence of the second coordination sphere on O2 activation by a nonheme iron(II) thiolate complex.J Inorg Biochem. 2025 Mar;264:112776. doi: 10.1016/j.jinorgbio.2024.112776. Epub 2024 Nov 17. J Inorg Biochem. 2025. PMID: 39644805
-
Spectroscopic, electrochemical, and kinetic trends in Fe(III)-thiolate disproportionation near physiologic pH.J Biol Inorg Chem. 2024 Apr;29(3):291-301. doi: 10.1007/s00775-024-02051-3. Epub 2024 May 9. J Biol Inorg Chem. 2024. PMID: 38722396 Free PMC article.
-
Destined for destruction: The role of methionine aminopeptidases and plant cysteine oxidases in N-degron formation.Plant Physiol. 2024 Dec 23;197(1):kiae667. doi: 10.1093/plphys/kiae667. Plant Physiol. 2024. PMID: 39875105 Free PMC article. Review.
-
Spectroscopic definition of ferrous active sites in non-heme iron enzymes.Methods Enzymol. 2024;703:29-49. doi: 10.1016/bs.mie.2024.05.019. Epub 2024 Jun 21. Methods Enzymol. 2024. PMID: 39261000 Free PMC article.
References
-
- Gardner JD; Pierce BS; Fox BG; Brunold TC Spectroscopic and Computational Characterization of Substrate-Bound Mouse Cysteine Dioxygenase: Nature of the Ferrous and Ferric Cysteine Adducts and Mechanistic Implications. Biochemistry 2010, 49 (29), 6033–6041. 10.1021/bi100189h. - DOI - PMC - PubMed
-
Resting, substrate cysteine (Cys), and substrate analogue selenocysteine (Sec) bound forms of CDO in the Fe(II) and Fe(III) states were investigated spectroscopically. Despite the identical binding modes of Cys and Sec, CDO was found to be unable to oxidize Sec.
-
- Blaesi EJ; Gardner JD; Fox BG; Brunold TC Spectroscopic and Computational Characterization of the NO Adduct of Substrate-Bound Fe(II) Cysteine Dioxygenase: Insights into the Mechanism of O2 Activation. Biochemistry 2013, 52 (35), 6040–6051. 10.1021/bi400825c. - DOI - PMC - PubMed
-
To explore the origin of the differences in reactivity between Cys- and Sec-bound CDO, the geometries and energies of viable O2-bound intermediates were evaluated computationally. A low-energy Fe(III)-superoxo intermediate could only be identified for Cys-bound CDO.
-
- Fernandez RL; Dillon SL; Stipanuk MH; Fox BG; Brunold TC Spectroscopic Investigation of Cysteamine Dioxygenase. Biochemistry 2020, 59 (26), 2450–2458. 10.1021/acs.biochem.0c00267. - DOI - PMC - PubMed
-
Magnetic circular dichroism and electron paramagnetic resonance data obtained for substrate and substrate analogue bound ADO provided compelling evidence that ADO features a 3-histidine facial triad, like cysteine dioxygenase, and revealed monodentate coordination of cysteamine to the Fe cofactor.
-
- Fernandez RL; Elmendorf LD; Smith RW; Bingman CA; Fox BG; Brunold TC The Crystal Structure of Cysteamine Dioxygenase Reveals the Origin of the Large Substrate Scope of This Vital Mammalian Enzyme. Biochemistry 2021, 60 (48), 3728–3737. 10.1021/acs.biochem.1c00463. - DOI - PMC - PubMed
-
The X-ray crystal structure of cysteamine dioxygenase revealed that the three-histidine active site is situated at the end of a wide substrate access channel, as in plant cysteine dioxygenase. Models of substrate bound ADO were generated computationally.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
