

A single introduction of wild rabbits triggered the biological invasion of Australia
- PMID: 35994668
- PMCID: PMC9436340
- DOI: 10.1073/pnas.2122734119
A single introduction of wild rabbits triggered the biological invasion of Australia
Abstract
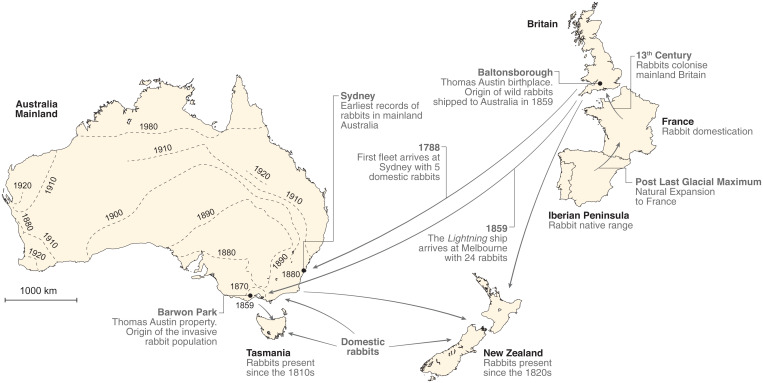
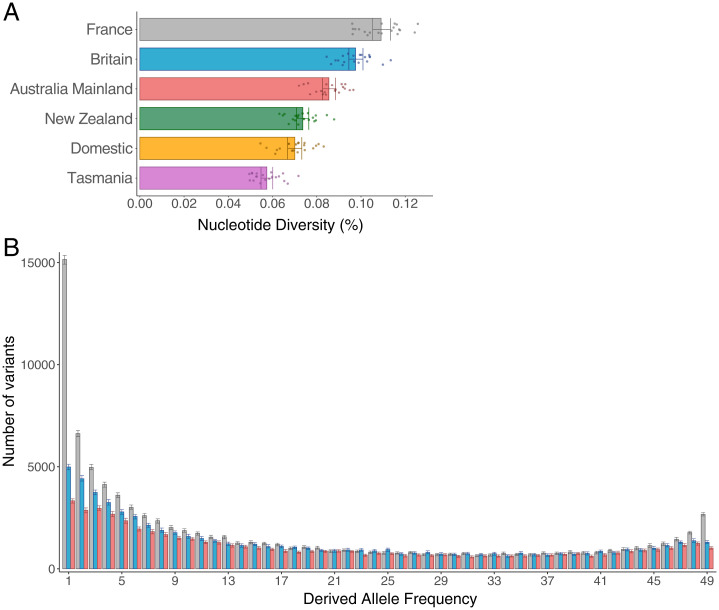
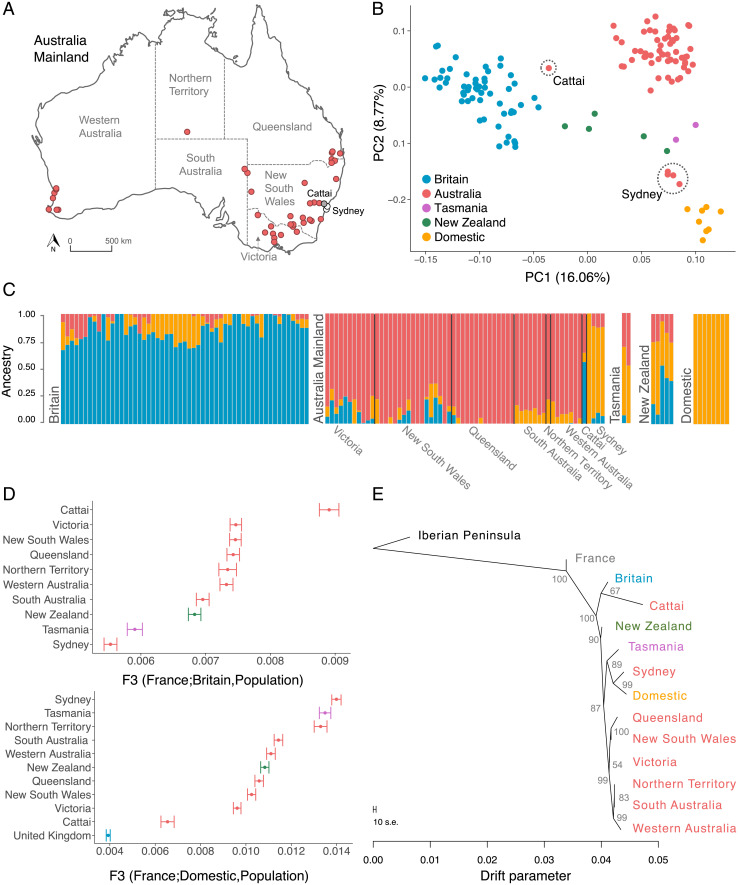
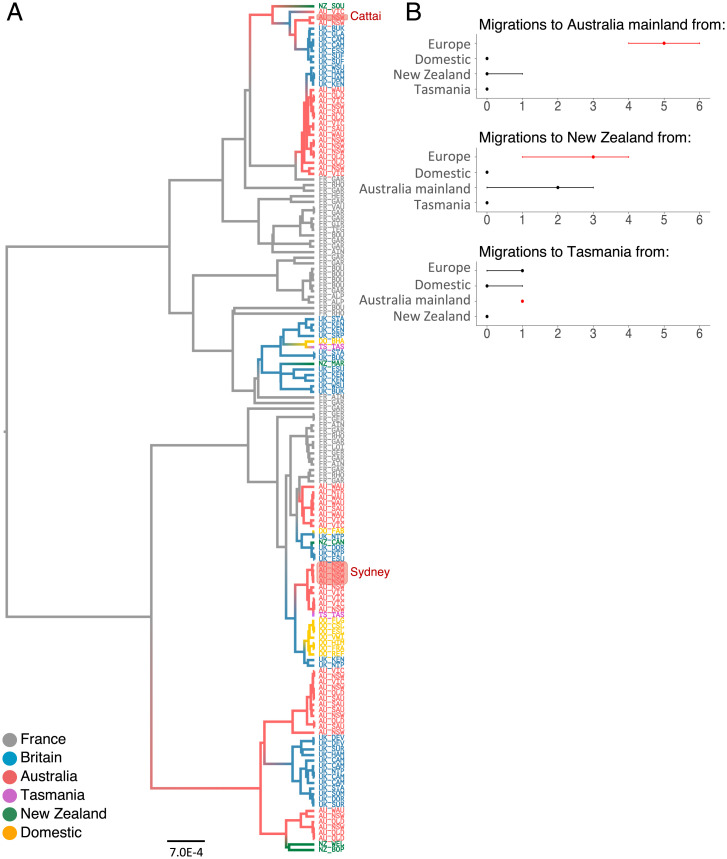
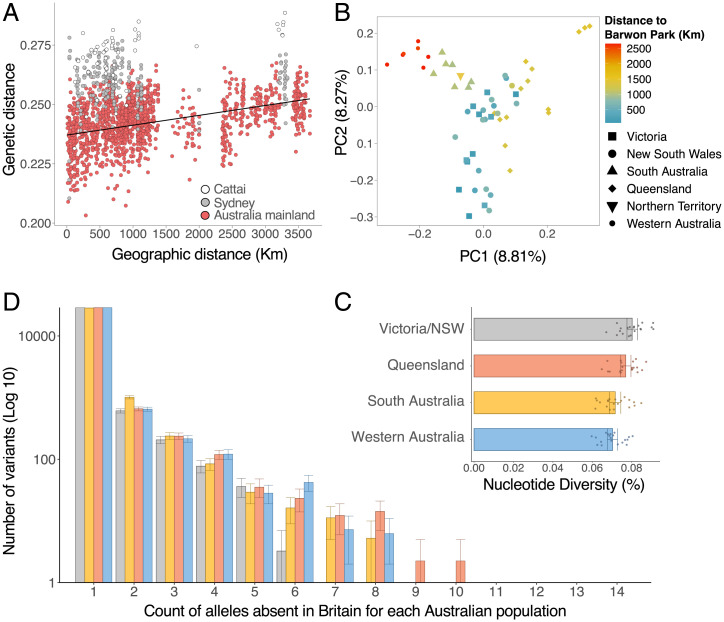
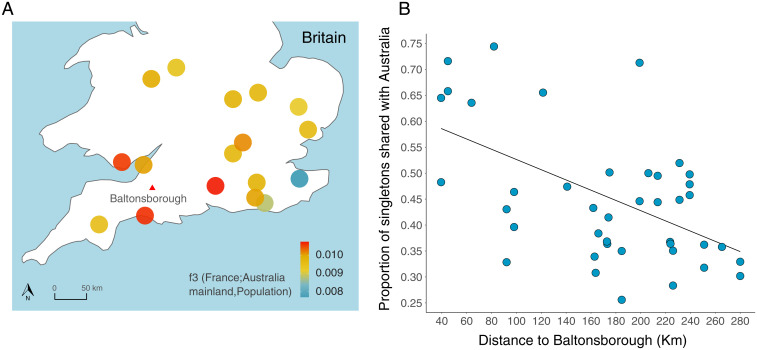
Biological invasions are a major cause of environmental and economic disruption. While ecological factors are key determinants of their success, the role of genetics has been more challenging to demonstrate. The colonization of Australia by the European rabbit is one of the most iconic and devastating biological invasions in recorded history. Here, we show that despite numerous introductions over a 70-y period, this invasion was triggered by a single release of a few animals that spread thousands of kilometers across the continent. We found genetic support for historical accounts that these were English rabbits imported in 1859 by a settler named Thomas Austin and traced the origin of the invasive population back to his birthplace in England. We also find evidence of additional introductions that established local populations but have not spread geographically. Combining genomic and historical data we show that, contrary to the earlier introductions, which consisted mostly of domestic animals, the invasive rabbits had wild ancestry. In New Zealand and Tasmania, rabbits also became a pest several decades after being introduced. We argue that the common denominator of these invasions was the arrival of a new genotype that was better adapted to the natural environment. These findings demonstrate how the genetic composition of invasive individuals can determine the success of an introduction and provide a mechanism by which multiple introductions can be required for a biological invasion.
Keywords: allele surfing; exome sequencing; invasion biology; population expansion.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Ehrenfeld J. G., Ecosystem consequences of biological invasions. Annu. Rev. Ecol. Evol. Syst. 41, 59–80 (2010).
-
- Diagne C., et al. , High and rising economic costs of biological invasions worldwide. Nature 592, 571–576 (2021). - PubMed
-
- Zenni R. D., Nuñez M. A., The elephant in the room: The role of failed invasions in understanding invasion biology. Oikos 122, 801–815 (2013).
-
- Simberloff D., The role of propagule pressure in biological invasions. Annu. Rev. Ecol. Evol. Syst. 40, 81–102 (2009).
-
- Lau J. A., terHorst C. P., Causes and consequences of failed adaptation to biological invasions: The role of ecological constraints. Mol. Ecol. 24, 1987–1998 (2015). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
