

The transcription factor Tbx5 regulates direction-selective retinal ganglion cell development and image stabilization
- PMID: 35998637
- PMCID: PMC9560999
- DOI: 10.1016/j.cub.2022.07.064
The transcription factor Tbx5 regulates direction-selective retinal ganglion cell development and image stabilization
Abstract
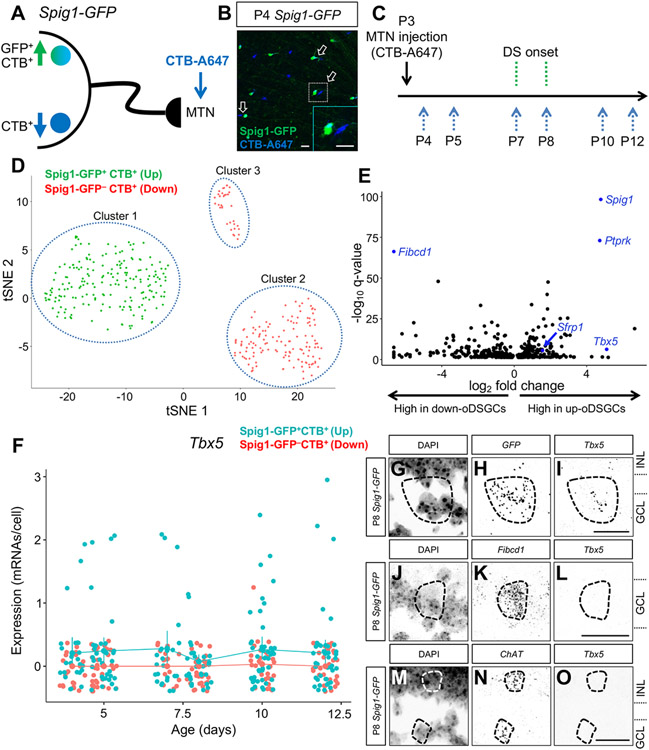
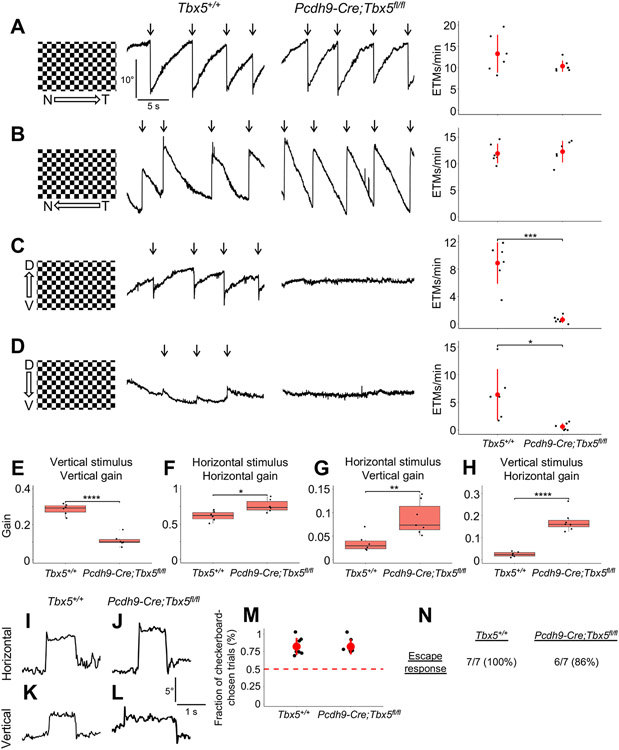
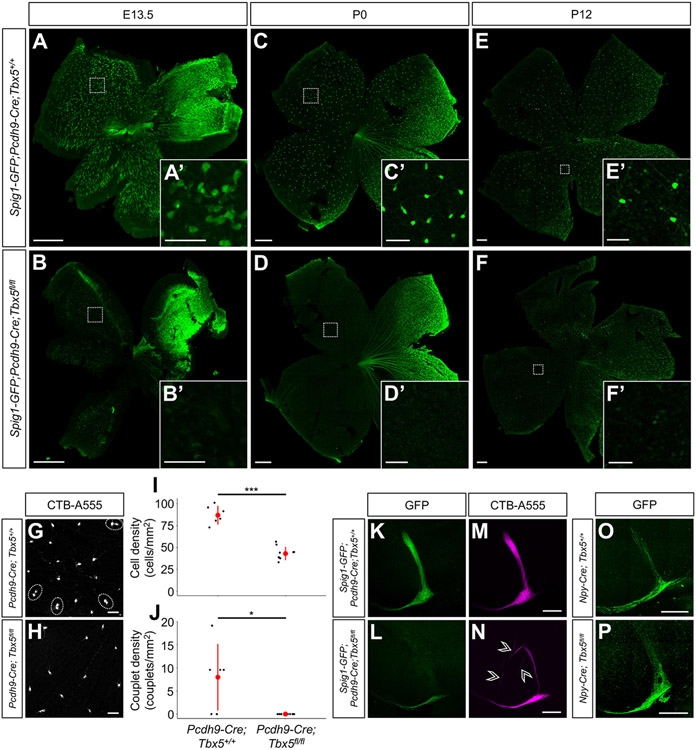
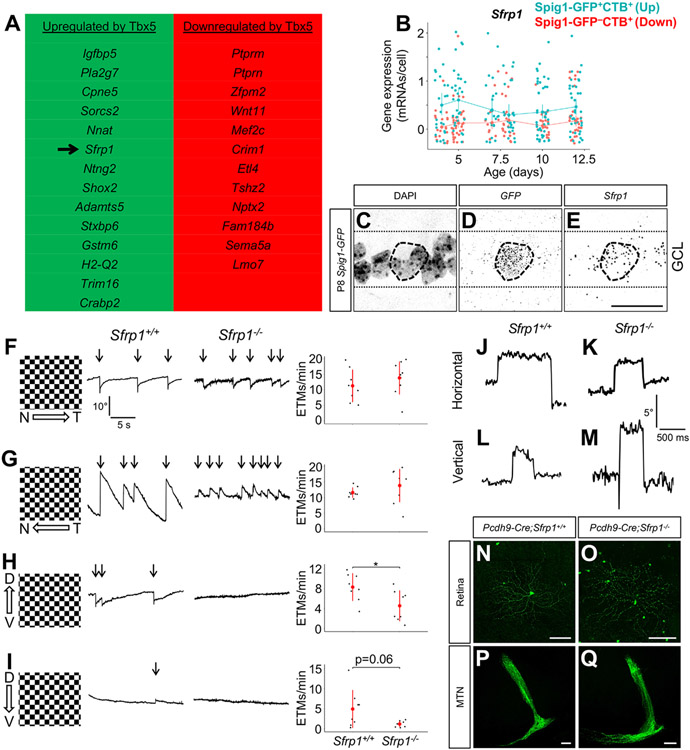
The diversity of visual input processed by the mammalian visual system requires the generation of many distinct retinal ganglion cell (RGC) types, each tuned to a particular feature. The molecular code needed to generate this cell-type diversity is poorly understood. Here, we focus on the molecules needed to specify one type of retinal cell: the upward-preferring ON direction-selective ganglion cell (up-oDSGC) of the mouse visual system. Single-cell transcriptomic profiling of up- and down-oDSGCs shows that the transcription factor Tbx5 is selectively expressed in up-oDSGCs. The loss of Tbx5 in up-oDSGCs results in a selective defect in the formation of up-oDSGCs and a corresponding inability to detect vertical motion. A downstream effector of Tbx5, Sfrp1, is also critical for vertical motion detection but not up-oDSGC formation. These results advance our understanding of the molecular mechanisms that specify a rare retinal cell type and show how disrupting this specification leads to a corresponding defect in neural circuitry and behavior.
Keywords: direction-selectivity; eye movements; neural development; retina; retinal ganglion cell; single-cell RNA-seq.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
References
-
- Tran NM, Shekhar K, Whitney IE, Jacobi A, Benhar I, Hong G, Yan W, Adiconis X, Arnold ME, Lee JM, Levin JZ, Lin D, Wang C, Lieber CM, Regev A, He Z, and Sanes JR (2019). Single-cell profiles of retinal ganglion cells differing in resilience to injury reveal neuroprotective genes. Neuron 104, 1039–1055.e12. 10.1016/j.neuron.2019.11.006. - DOI - PMC - PubMed
-
- Brodie-Kommit J, Clark BS, Shi Q, Shiau F, Kim DW, Langel J, Sheely C, Ruzycki PA, Fries M, Javed A, Cayouette M, Schmidt T, Badea T, Glaser T, Zhao H, Singer J, Blackshaw S, and Hattar S (2021). Atoh7-independent specification of retinal ganglion cell identity. Sci. Adv 7, eabe4983. 10.1126/sciadv.abe4983. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
