

An amphipathic helix in Brl1 is required for nuclear pore complex biogenesis in S. cerevisiae
- PMID: 36000978
- PMCID: PMC9402233
- DOI: 10.7554/eLife.78385
An amphipathic helix in Brl1 is required for nuclear pore complex biogenesis in S. cerevisiae
Abstract
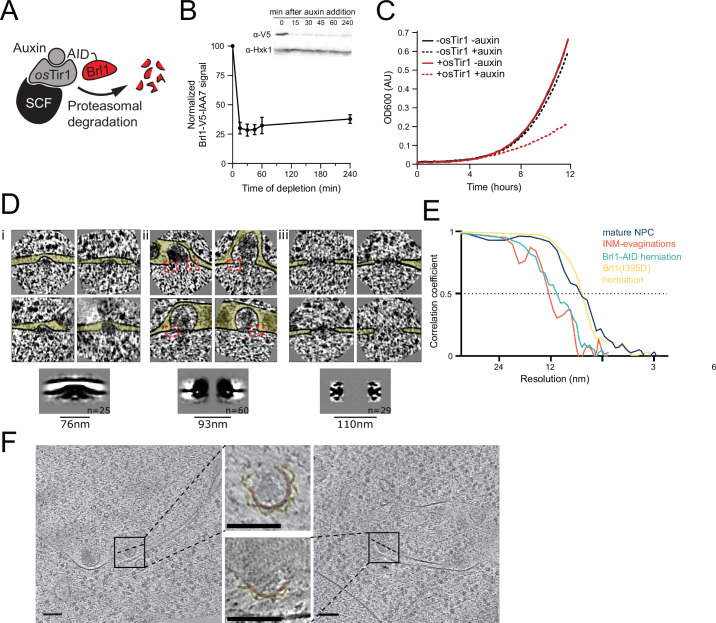
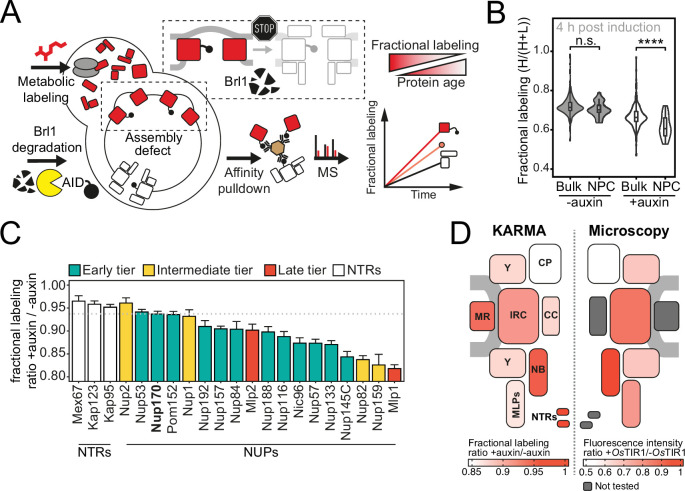
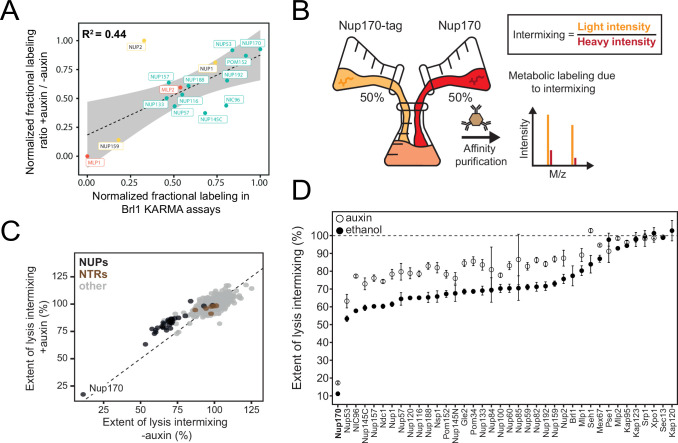
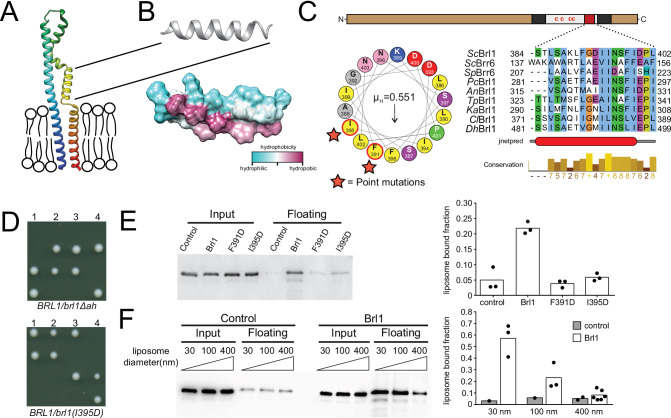
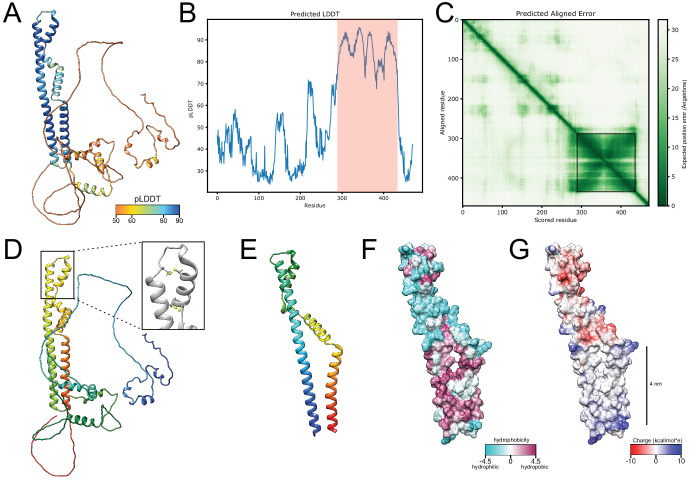
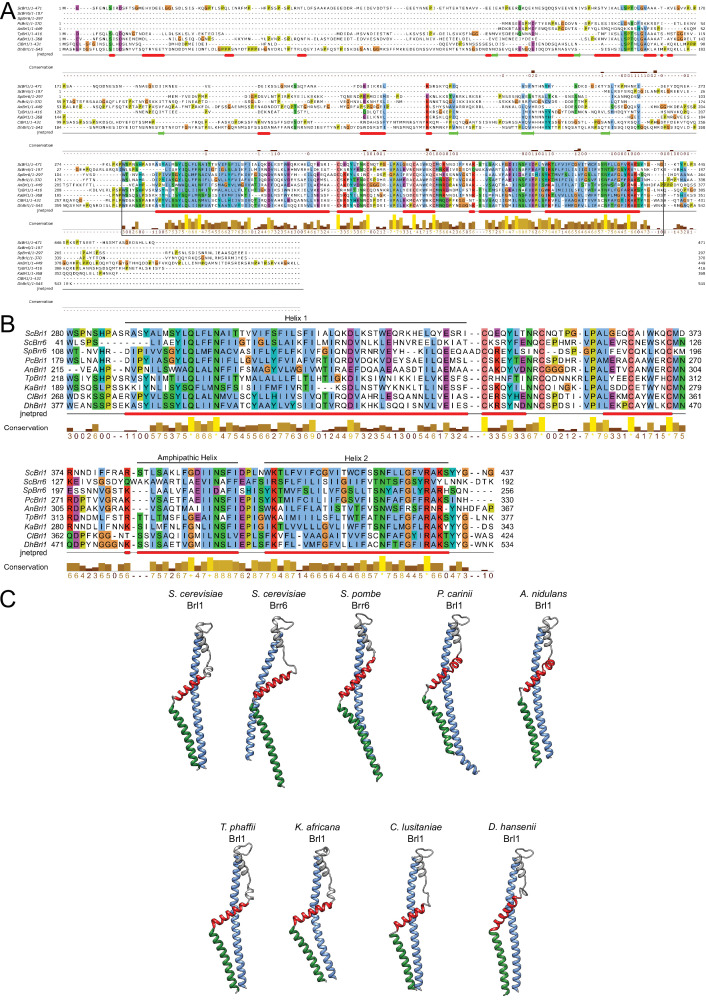
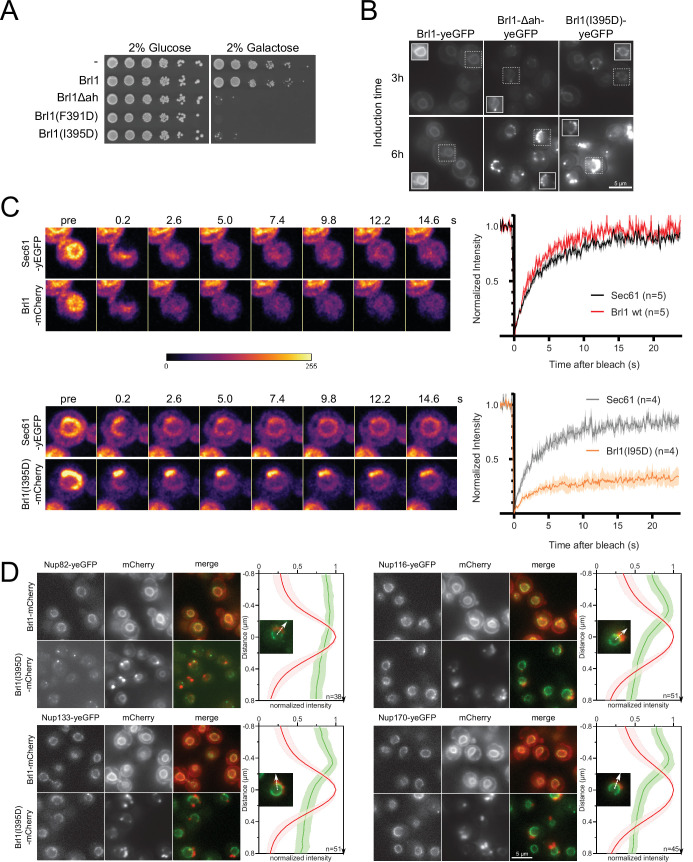
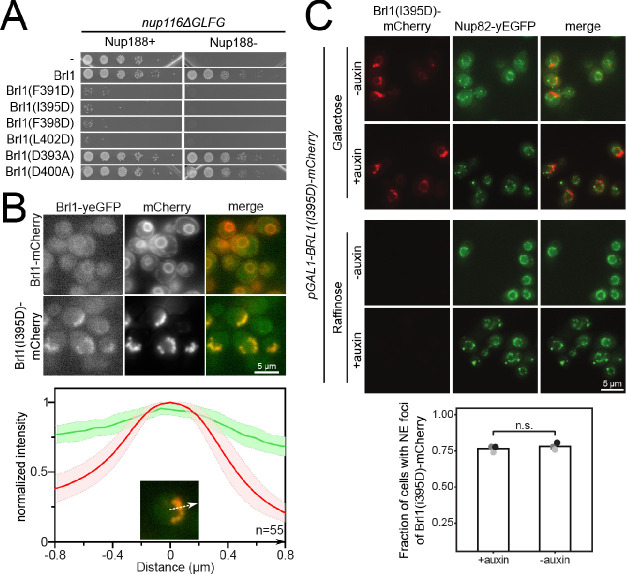
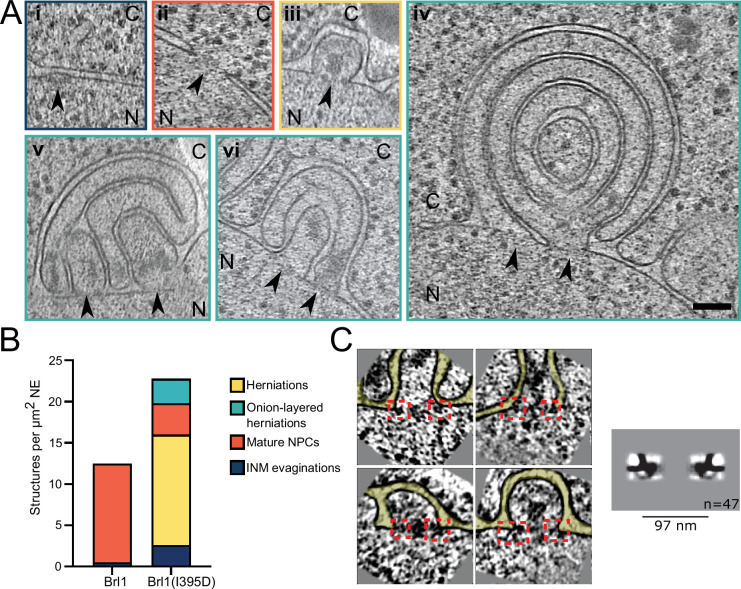
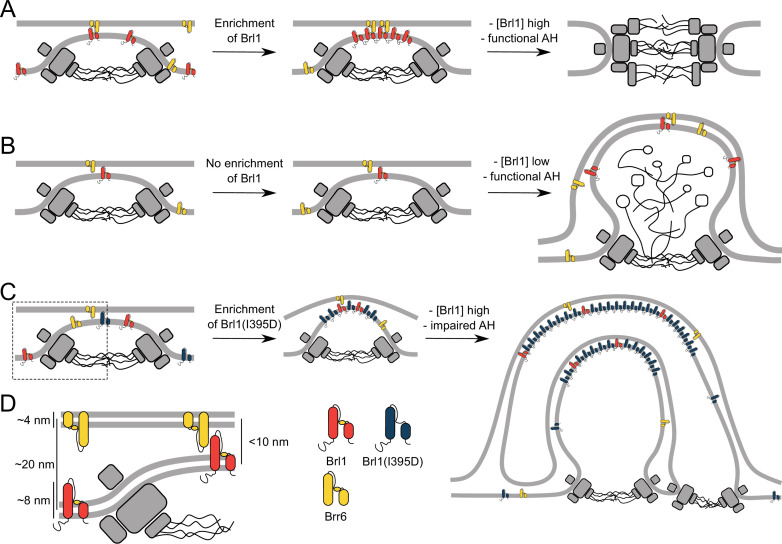
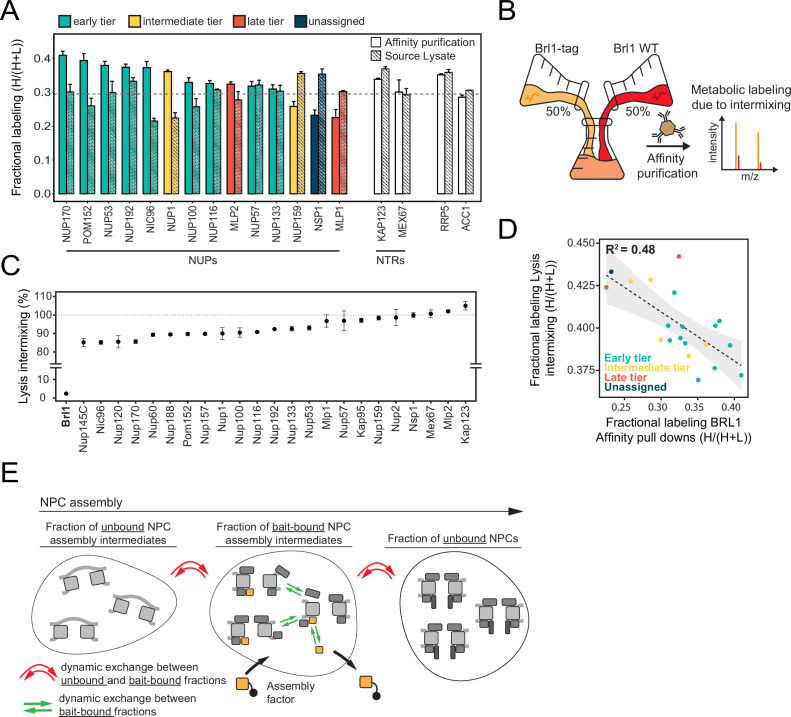
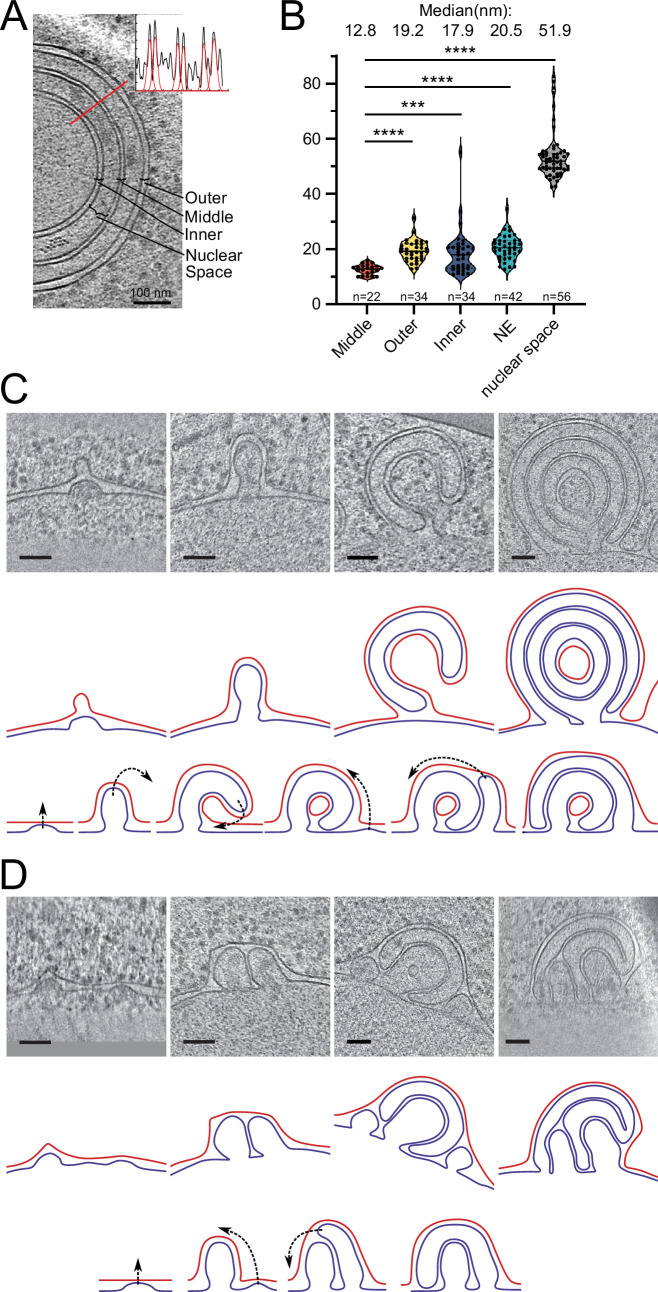
The nuclear pore complex (NPC) is the central portal for macromolecular exchange between the nucleus and cytoplasm. In all eukaryotes, NPCs assemble into an intact nuclear envelope (NE) during interphase, but the process of NPC biogenesis remains poorly characterized. Furthermore, little is known about how NPC assembly leads to the fusion of the outer and inner NE, and no factors have been identified that could trigger this event. Here, we characterize the transmembrane protein Brl1 as an NPC assembly factor required for NE fusion in budding yeast. Brl1 preferentially associates with NPC assembly intermediates and its depletion halts NPC biogenesis, leading to NE herniations that contain inner and outer ring nucleoporins but lack the cytoplasmic export platform. Furthermore, we identify an essential amphipathic helix in the luminal domain of Brl1 that mediates interactions with lipid bilayers. Mutations in this amphipathic helix lead to NPC assembly defects, and cryo-electron tomography analyses reveal multilayered herniations of the inner nuclear membrane with NPC-like structures at the neck, indicating a failure in NE fusion. Taken together, our results identify a role for Brl1 in NPC assembly and suggest a function of its amphipathic helix in mediating the fusion of the inner and outer nuclear membranes.
Keywords: S. cerevisiae; cell biology; membrane fusion; nuclear envelope; nuclear pore complex.
© 2022, Kralt, Wojtynek, Fischer et al.
Conflict of interest statement
AK, MW, JF, AA, RM, ED, EN, FU, MT, WA, EO, OM, KW No competing interests declared
Figures
Similar articles
-
The Nuclear Pore Complex: Birth, Life, and Death of a Cellular Behemoth.Cells. 2022 Apr 25;11(9):1456. doi: 10.3390/cells11091456. Cells. 2022. PMID: 35563762 Free PMC article. Review.
-
A perinuclear α-helix with amphipathic features in Brl1 promotes NPC assembly.Mol Biol Cell. 2022 May 1;33(5):ar35. doi: 10.1091/mbc.E21-12-0616. Epub 2022 Mar 16. Mol Biol Cell. 2022. PMID: 35293775 Free PMC article.
-
Brr6 and Brl1 locate to nuclear pore complex assembly sites to promote their biogenesis.J Cell Biol. 2018 Mar 5;217(3):877-894. doi: 10.1083/jcb.201706024. Epub 2018 Feb 9. J Cell Biol. 2018. PMID: 29439116 Free PMC article.
-
A short perinuclear amphipathic α-helix in Apq12 promotes nuclear pore complex biogenesis.Open Biol. 2021 Nov;11(11):210250. doi: 10.1098/rsob.210250. Epub 2021 Nov 24. Open Biol. 2021. PMID: 34814743 Free PMC article.
-
Nuclear pore biogenesis into an intact nuclear envelope.Chromosoma. 2010 Oct;119(5):469-77. doi: 10.1007/s00412-010-0289-2. Epub 2010 Aug 19. Chromosoma. 2010. PMID: 20721671 Review.
Cited by
-
Nuclear basket proteins regulate the distribution and mobility of nuclear pore complexes in budding yeast.Mol Biol Cell. 2024 Nov 1;35(11):ar143. doi: 10.1091/mbc.E24-08-0371. Epub 2024 Sep 25. Mol Biol Cell. 2024. PMID: 39320946 Free PMC article.
-
Lipid saturation controls nuclear envelope function.Nat Cell Biol. 2023 Sep;25(9):1290-1302. doi: 10.1038/s41556-023-01207-8. Epub 2023 Aug 17. Nat Cell Biol. 2023. PMID: 37591950 Free PMC article.
-
The Nuclear Pore Complex: Birth, Life, and Death of a Cellular Behemoth.Cells. 2022 Apr 25;11(9):1456. doi: 10.3390/cells11091456. Cells. 2022. PMID: 35563762 Free PMC article. Review.
-
Elucidating the nanoscopic organization and dynamics of the nuclear pore complex.Nucleus. 2025 Dec;16(1):2510106. doi: 10.1080/19491034.2025.2510106. Epub 2025 Jun 4. Nucleus. 2025. PMID: 40464641 Free PMC article.
-
CLCC1 promotes hepatic neutral lipid flux and nuclear pore complex assembly.bioRxiv [Preprint]. 2024 Jun 8:2024.06.07.597858. doi: 10.1101/2024.06.07.597858. bioRxiv. 2024. PMID: 38895340 Free PMC article. Preprint.
References
-
- Akey CW, Singh D, Ouch C, Echeverria I, Nudelman I, Varberg JM, Yu Z, Fang F, Shi Y, Wang J, Salzberg D, Song K, Xu C, Gumbart JC, Suslov S, Unruh J, Jaspersen SL, Chait BT, Sali A, Fernandez-Martinez J, Ludtke SJ, Villa E, Rout MP. Comprehensive structure and functional adaptations of the yeast nuclear pore complex. Cell. 2022;185:361–378. doi: 10.1016/j.cell.2021.12.015. - DOI - PMC - PubMed
-
- Allegretti M, Zimmerli CE, Rantos V, Wilfling F, Ronchi P, Fung HKH, Lee C-W, Hagen W, Turoňová B, Karius K, Börmel M, Zhang X, Müller CW, Schwab Y, Mahamid J, Pfander B, Kosinski J, Beck M. In-cell architecture of the nuclear pore and snapshots of its turnover. Nature. 2020;586:796–800. doi: 10.1038/s41586-020-2670-5. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
