

Glucagon receptor signaling at white adipose tissue does not regulate lipolysis
- PMID: 36002172
- PMCID: PMC9576180
- DOI: 10.1152/ajpendo.00078.2022
Glucagon receptor signaling at white adipose tissue does not regulate lipolysis
Abstract
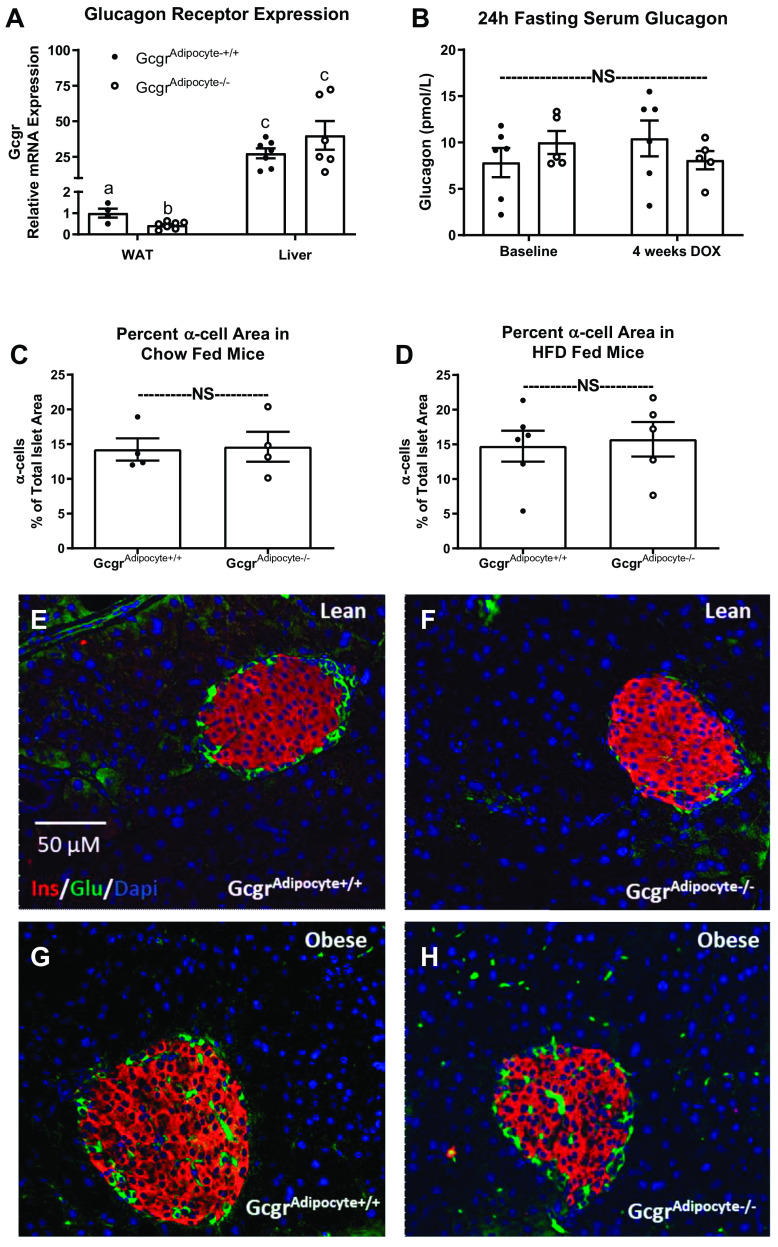
Although the physiological role of glucagon receptor signaling in the liver is well defined, the impact of glucagon receptor (Gcgr) signaling on white adipose tissue (WAT) continues to be debated. Although numerous studies propose that glucagon stimulates WAT lipolysis, we lack evidence that physiological concentrations of glucagon regulate WAT lipolysis. In turn, we performed studies in both wild-type and WAT Gcgr knockout mice to determine if glucagon regulates lipolysis at WAT in the mouse. We assessed the effects of fasting and acute exogenous glucagon administration in wild-type C57BL/6J and GcgrAdipocyte+/+ versus GcgrAdipocyte-/- mice. Using an ex vivo lipolysis protocol, we further examined the direct effects of glucagon on physiologically (fasted) and pharmacologically stimulated lipolysis. We found that adipocyte Gcgr expression did not affect fasting-induced lipolysis or hepatic lipid accumulation in lean or diet-induced obese (DIO) mice. Acute glucagon administration did not affect serum nonesterified fatty acids (NEFA), leptin, or adiponectin concentration, but did increase serum glucose and FGF21, regardless of genotype. Glucagon did not affect ex vivo lipolysis in explants from either GcgrAdipocyte+/+ or GcgrAdipocyte-/- mice. Gcgr expression did not affect fasting-induced or isoproterenol-stimulated lipolysis from WAT explants. Moreover, glucagon receptor signaling at WAT did not affect body weight or glucose homeostasis in lean or DIO mice. Our studies have established that physiological levels of glucagon do not regulate WAT lipolysis, either directly or indirectly. Given that glucagon receptor agonism can improve dyslipidemia and decrease hepatic lipid accumulation, it is critical to understand the tissue-specific effects of glucagon receptor action. Unlike the crucial role of hepatic glucagon receptor signaling in maintaining glucose and lipid homeostasis, we observed no metabolic consequence of WAT glucagon receptor deletion.NEW & NOTEWORTHY It has been postulated that glucagon stimulates lipolysis and fatty acid release from white adipose tissue. We observed no metabolic effects of eliminating or activating glucagon receptor signaling at white adipose tissue.
Keywords: fasting; glucagon; lipolysis; type 2 diabetes; white adipose tissue.
Conflict of interest statement
No conflicts of interest, financial or otherwise, are declared by the authors.
Figures





References
-
- Longuet C, Robledo AM, Dean ED, Dai C, Ali S, McGuinness I, de Chavez V, Vuguin PM, Charron MJ, Powers AC, Drucker DJ. Liver-specific disruption of the murine glucagon receptor produces α-cell hyperplasia: evidence for a circulating α-cell growth factor. Diabetes 62: 1196–1205, 2013. doi: 10.2337/db11-1605. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases